Biochar: the Black 'Magic' of Terra Preta
How a high-surface-area carbon lattice may drive soil microbial communities across a threshold into a self-stabilizing, high-fertility state, and why terra preta is grown rather than manufactured
Abstract
Amazonian dark earth (terra preta) is more fertile, more biologically active, and more persistent than the weathered tropical soils it overlies, holding fertility for centuries after abandonment. This paper advances a single mechanistic hypothesis and the established science motivating it: that a durable, high-surface-area pyrogenic carbon lattice acts as a multifunctional shared substrate which, supplied together with comprehensive mineral resources and a microbial inoculum, lets ordinary soil self-organize under selection into a qualitatively different state of ecological organization — its endpoint shaped by founding conditions and local biota, not fixed. The argument has four parts. First, the carbon lattice supplies habitable space, ion-exchange capacity, nutrient and water reservoirs, signal retention, and shelter, through pore-size classes and surface chemistries that mature on different timescales. Second, the resulting rise in microbial density and proximity switches on a community economy governed by quorum sensing and metabolic division of labor; across an evolutionary clock, the Black Queen dynamic of adaptive gene loss compounds that economy, deepening interdependence and stability. Third, the positive feedbacks of that economy are the formal prerequisite for an alternative stable state, and soil microbial communities are increasingly inferred to occupy alternative stable states — the rigorous form of an ecological “escape velocity,” a tipping point into a self-reinforcing basin. Fourth, fungi integrate the system, mobilizing minerals, trading reciprocally with hosts, and priming immunity, with these functions scaling with mycelial density. On this account, persistence is itself two coupled mechanisms of unlike kind: a kinetically stable carbon scaffold that endures passively, and an actively self-reinforcing community that holds fertility only while fed — the scaffold a durable home for the living engine, which supplies what the scaffold cannot. The same organization predicts a distinctive efficiency profile — tight internal nutrient cycling, retention-driven water economy, and self-replenishing cycling under continued cropping — as an emergent consequence. Each component is independently established across five disciplines; their convergence on one architecture is what makes the hypothesis worth testing, though convergence is not assembly in soil. What would convert it into a discovery is one experiment: an engineered up-transition with hysteresis in real soil.
1. Introduction: terra preta as a grown soil
Amazonian dark earth has long carried a reputation bordering on the magical: a black soil that turns barren tropical ground productive and holds it that way for generations. This paper argues that the "magic" resolves into mechanism, and that among the engines of that mechanism is a named evolutionary dynamic, the Black Queen, from which this paper takes part of its title. The reputation is earned; the explanation is ordinary science.
The central puzzle of terra preta is not that it is fertile. Many soils are fertile while inputs continue. The puzzle is that terra preta remained fertile, biologically rich, and structurally distinct for centuries to millennia after deliberate inputs ceased, sitting in the middle of a landscape of strongly weathered, nutrient-poor Ferralsols and Acrisols that do not behave that way. A soil that holds a high-function state without ongoing maintenance is behaving less like a fertilized field and more like a system that has settled into a different equilibrium.
The hypothesis advanced here is that this difference is produced, and then maintained, by a physical object: a stable, high-surface-area carbon lattice derived from pyrolyzed biomass. Supplied alongside a complete mineral profile (macronutrients and a comprehensive complement of trace elements) and a microbial inoculum, the lattice does one specific and consequential thing. It adds an enormous quantity of structured habitat and reactive surface. That addition lifts microbial density and, more importantly, microbial proximity, to the point where the community is no longer limited by space, by the diffusion of shared resources, or by the loss of metabolic partners. Past a threshold, with resources in surplus so that nothing is rate-limiting, the system does not simply hold more of the same life. It crosses into a different regime of ecological organization that can both assemble and remain assembled, and it is that self-stabilizing, higher-function state that this paper identifies with terra preta.
The claim of this paper can be stated as a single proposition: the conditioned carbon lattice is the keystone variable of the terra preta syndrome. One material, charcoal of sufficient quantity, matured to high exchange capacity, loaded with a complete mineral profile, and inoculated, is the controlling input from which the remaining properties of the soil follow, rather than a list of independently supplied traits that must each be installed. Those properties resolve into two dividends on two clocks. The first is immediate and physical: porosity and tilth, water retention, and the resistance to leaching that a high-surface-area, high-exchange substrate confers the moment it enters the ground, requiring no biology and no maturation. The second is slower and biological: the self-replenishing fertility that persists under cropping, the suppression of disease, and the organized microbial complexity that emerge as ordinary soil physics and biology operate on that substrate over time. The strong form of the proposition is that this second dividend is genuinely emergent, the outcome of feedbacks running on a suitable substrate rather than a set of separately engineered functions, and that it is therefore reproducible in principle by conditioning carbon to the same specification, which reframes the common view that terra preta cannot be deliberately replicated as a tractable problem of substrate conditioning rather than an irreducible mystery. The proposition is bounded in one respect the body develops: the biological dividend is reliably emergent under adequate inoculation and sufficient time, not instantaneous and not indifferent to founding conditions, because priority effects in community assembly mean the same substrate can be colonized along more than one trajectory.
Three points of precision are worth stating at the outset, because they distinguish a defensible claim from an overstated one.
First, the transition is best described as a tipping point into an alternative basin of attraction, not as a literal escape to an unbounded state. The community does not run away; it settles into a higher, self-reinforcing equilibrium that resists perturbation. This framing, developed in Section 4, is what makes the persistence of terra preta a feature of the mechanism rather than a coincidence.
Second, the carbon lattice is proposed as an unusually efficient and durable route to high-surface-area structured habitat, not as the only possible route. Chernozems and deep compost-built soils reach high density and complexity with little pyrogenic carbon. Pyrolytic char is distinguished by the durability of its physical structure and by the breadth of functions its surface supports simultaneously, not by being uniquely necessary. Within the Amazonian comparison, however, the lattice is the keystone difference. Terra preta differs from its neighbors in mineral nutrients and in biology as well, but those can be added to ordinary soils that then lose them again; what the dark earth additionally has, and what makes its added fertility persist, is the durable carbon lattice that holds and organizes those minerals and that biology in place. Keystone here denotes the element whose removal would collapse the persistent result, not the only element present.
Third, the claim is mechanistically grounded but not yet demonstrated as a controlled regime shift in real soil. The synthesis that follows shows that every component the mechanism requires is independently well established. The integrated outcome, an engineered up-transition into a self-stabilizing high-fertility state, remains the open experiment, and the paper is explicit about where that boundary lies.
2. The carbon lattice as a multifunctional shared substrate
The starting observation is quantitative. Pyrolytic char can present a very large specific surface area, with reported values across the literature spanning roughly twenty to several hundred square meters per gram for plant-derived chars, and higher still for activated carbons. The soils into which such char is most relevant are, by contrast, low-surface-area materials. Sand is effectively near zero; silt is on the order of one square meter per gram; and kaolinite, the non-swelling one-to-one clay that dominates the strongly weathered tropical soils of the Amazonian comparison, sits at roughly ten to thirty square meters per gram. The implication is that adding even a modest fraction of char to a low-surface-area soil can raise total surface area substantially. For a soil with a genuinely low clay baseline, a char-amended blend can approach an order-of-magnitude increase in total surface area; for higher baselines and lower-surface-area chars the gain is smaller, on the order of several-fold. The defensible statement is therefore "a several-fold to roughly order-of-magnitude increase in total surface area, reaching a full order of magnitude in sandy, weathered, low-clay soils," which is precisely the soil context in which terra preta arose.
The important refinement is that "surface area," as measured by gas adsorption, is not equivalent to habitable space for a microorganism. Gas-adsorption surface area is dominated by micropores far too small for cells to occupy. The fraction that is open and accessible is considerably smaller than the total, and the pores that actually house microbes are micron-scale macropores, broadly in the range of a few to roughly twenty micrometers, consistent with cell dimensions of one to five micrometers and with fungal hyphal colonization of larger pores. A char surface-area figure therefore overstates colonizable habitat, often by several-fold.
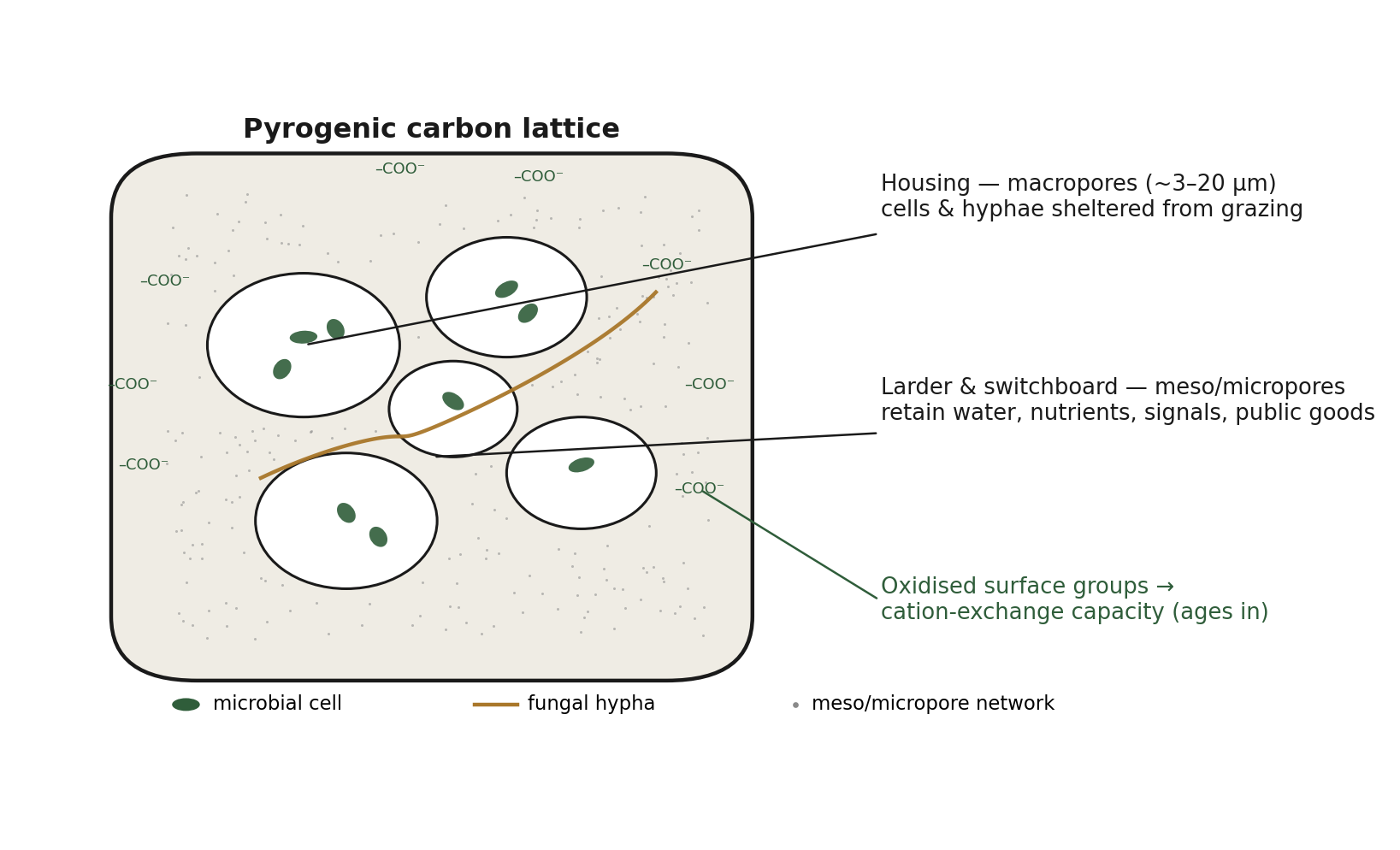
This distinction does not weaken the thesis; it sharpens it into a two-channel model in which the same material does different jobs through different pore-size classes (Figure 1):
Housing. The micron-scale macropores are where cells and hyphae live, and where they are physically sheltered from grazing protozoa. This shelter is a mechanism for realized density gains, not merely potential ones.
Larder and switchboard. The finer mesoporous and microporous surface, the bulk of the gas-adsorption area, adsorbs and retains water, dissolved organic matter, gases, mineral nutrients, and the diffusible signals and public goods of the microbial community, holding them close instead of allowing them to wash away.
The second channel is the physical basis for the community economy described in Section 3, and the coupling of the two channels through one piece of material is the heart of the "multifunctional substrate" idea.
A further and frequently misstated point concerns cation exchange capacity (CEC), the soil's capacity to hold exchangeable nutrient cations. CEC does not scale with bare surface area. It arises from oxygen-bearing functional groups (carboxyl, phenolic, and hydroxyl groups) on the carbon surface. Because high pyrolysis temperature develops surface area and porosity while removing those functional groups, surface area and fresh CEC tend to anti-correlate across temperature, with CEC peaking at relatively low pyrolysis temperatures. This appears at first to be a problem for any "more surface, more function" claim, but two considerations resolve it and, in doing so, strengthen the maturation thesis.
The first consideration is that habitat, surface area, and CEC are set by three different variables, and are therefore partly independent design levers rather than a single slider. Macroporous habitat derives largely from the preserved cellular architecture of the feedstock; gas-adsorption surface area derives from high-temperature carbonization; CEC derives from low-temperature and, importantly, aged surface chemistry. Because application rate is a free additional lever, there is no obligatory trade of CEC against surface area at the system level: a higher dose of a lower-temperature, higher-CEC char can supply both habitat and exchange capacity, at the cost of more material rather than a loss of function.
The second consideration is temporal, and it is the more interesting one. The physical channel (surface area and pore structure) is fixed at the moment of pyrolysis, whereas much of the surface chemistry (and therefore much of the CEC and a large part of the biological function) develops over time as the char weathers and its surface oxidizes, building carboxyl and carbonyl groups and negative charge. The housing is ready on day one; the ion-exchange economy and the associated biological function ripen over months to years. Different functions thus come online on different clocks. This asynchronous maturation is not a complication to be explained away. It is the mechanistic reason that terra preta is something grown rather than mixed: if every function switched on at once at the moment of application, there would be nothing to wait for, and the empirical observation that dark earth develops over time would have no basis.
A consequence of this exchange and sorption capacity is the inhibition of nutrient leaching, and it matters most in exactly the setting where terra preta arose. In a high-rainfall tropical landscape over kaolinitic, low-charge soils, soluble nutrients from rain and irrigation are flushed below the root zone before they can be taken up; the charged and sorptive lattice intercepts and holds those cations and dissolved organic compounds against that flux, so that fertility added to the soil stays in the soil rather than draining away. This anti-leaching role is one more retention function of the same surface, and a simple leaching column is among the most direct tests of it (Section 10).
The retention principle extends to water, through a buffer that operates on two levels. The first is physical: the pore structure adsorbs and holds water, raising the soil's water-holding capacity, most noticeably in coarse, low-clay soils. The second is biological: the dense community the lattice supports secretes extracellular polysaccharides and mucilage and builds the aggregate structure that together retain moisture and slow drying, so that a larger and more active biomass becomes, in effect, a second water reservoir built upon the first. The buffer against drought is therefore doubled, once by the physical cavities and again by the biology those cavities house. The water held in the biological term is held in the secreted matrix and the improved structure, not stored inside the cells themselves, whose volume is a negligible fraction of soil water; the mechanism is matrix and aggregation, not cellular storage.
These retention properties carry a corollary that is easy to miss and consequential for how the lattice should be prepared and tested. A fresh, uncharged char is not yet any of these things: its exchange sites and sorptive surfaces are empty, so on entering a soil it first draws nutrients and water out of solution and holds them, behaving for a time as a net sink that demands from its surroundings rather than a source that contributes to them. Only once those sites are loaded, the surface aged, and the biology established does the same material become a net contributor, releasing and cycling what it holds. This transition from sink to source is the practical face of the asynchronous maturation described above, and it is why the manner and the timing of preparation, and not the bare presence of char, determine whether the lattice helps or harms in its first seasons. It is also the most likely reason that short trials of fresh, uninoculated char report such inconsistent results, a point developed in Section 8, where this paper addresses the wider biochar literature.
The honest boundary of this section is that what is well established is the existence of large total surface area, the macropore limitation on habitable fraction, the housing-and-shelter role of those macropores, the retention of nutrients, water, and signals by the finer porosity, the resistance of that retention to leaching, and the initial net-sink behavior of a fresh, uncharged lattice. What is inferred, and addressed in Section 4, is that these properties together drive a discrete change in community organization.
3. The efficiency economy: density, division of labor, and reductive interdependence
If the carbon lattice raises density and proximity, the question becomes what changes when a microbial community becomes dense and tightly packed. The microbial-ecology literature answers this with several named, well-supported mechanisms that, taken together, describe a self-reinforcing economy.
Density-gated collective behavior. Quorum sensing is the canonical case of a microbial community switching behavior at a density threshold. Cells secrete signaling molecules whose concentration rises with population density; above a threshold concentration, the population activates collective functions such as biofilm formation, coordinated metabolism, and resource utilization (Miller and Bassler, 2001; Waters and Bassler, 2005). The claim that "past a certain density the system enters a new behavioral mode" is therefore not speculative; it is a central paradigm of bacterial sociobiology. One calibration is warranted: the sharpness of the threshold is contested, and single-cell studies indicate the switch can be graded rather than a hard cutoff. The defensible phrasing is "a threshold-like transition into a new regime," which is sufficient for the present argument.
Confinement lowers the density required. Beyond classical density sensing, there is strong support for diffusion sensing, in which quorum-sensing systems respond to physical containment, so that even an isolated cell in a sufficiently small, enclosed space can become effectively "quorate" because its signals do not diffuse away (Redfield, 2002). Interpreted in the context of a porous carbon lattice, a cell within a small pore behaves as though it is in a dense crowd. The structured habitat does not merely hold more cells; it lowers the effective density required to trigger collective behavior. This is a direct mechanistic link between pore space and the community economy.
Division of labor, and the proximity condition that unifies the argument. Distributing a long metabolic pathway across specialists reduces the per-cell burden relative to a single organism carrying the entire pathway, and modeling predicts that this can raise community productivity and allow more species to coexist on the same resources. The crucial and frequently omitted result is that division of labor is not automatically more efficient. Splitting a pathway across cells imposes a transport cost, because intermediates must move between populations, and a rigorous analysis shows that a two-strain consortium produces less biomass than an equivalent single organism of the same metabolic capacity unless specialization raises per-pathway efficiency enough to overcome that transport penalty (Tsoi et al., 2018). The variable that sets the transport penalty is distance. In a dilute, well-mixed system, intermediates leak away between partners and division of labor can lose; pack the partners micron-distances apart in structured habitat and the transport cost collapses, tilting the balance so that the efficiency economy wins. This is the point at which the two principal claims of this paper converge: proximity supplied by the carbon lattice is the variable that flips division of labor from a net cost into a net gain. The "real estate" claim and the "efficiency" claim are not two assertions but one mechanism.
The Black Queen dynamic: a second, evolutionary clock. The mechanisms above act on physiological time and need no genetic change, since densely packed cells simply perform them. A further dynamic acts on a different kind of clock, an evolutionary one that changes gene content, and rather than competing with the proximity economy it compounds upon it. The Black Queen Hypothesis (Morris, Lenski, and Zinser, 2012) holds that loss of a costly but leaky function is favored at the individual level and proceeds until production of that public good is just sufficient for the community, at which point further loss is checked by its cost. An organism gains by not spending resources to make what its neighbors already supply, which drives genome streamlining and produces obligate interdependence. Because this requires mutation and selection across generations, it is not what establishes the high-function state within a single brew cycle or a single season; it enters the system from two directions instead. First, the organisms that inoculate a soil are already auxotrophic and cross-dependent, because the shedding of biosynthetic genes is favored and pervasive in nature, common across both free-living and symbiotic bacteria, and the difficulty of culturing most soil microbes in isolation is a direct reflection of that dependence (D'Souza et al., 2014). The proximity economy therefore draws on a deep, ready-made web of interdependence from the first day, and the lattice supplies the dense, structured habitat in which those inherited dependencies can finally express themselves. Second, continued gene loss proceeds within the established community, and although it is fast when counted in generations, with auxotrophs evolving in fewer than two thousand generations under favorable conditions (D'Souza and Kost, 2016), it is paced in real time by the fact that most soil cells grow slowly, so it unfolds over the soil's maturation and its long life rather than within a season. As it proceeds, public goods are reliably supplied, selection on the now-redundant private copies relaxes, interdependence tightens, and the basin of attraction deepens. The dynamic does not establish the high-function state on its own; it makes a state already established by proximity progressively more entrenched and harder to dislodge as time passes. A modeled extension indicates that this process can be generative as well as reductive, with relaxed selection on redundant genes permitting the exploration of new functions, so that the economy compounds rather than merely plateaus (Takeuchi et al., 2024). The honest boundary is that this second clock, and especially its generative form, is an evolutionary argument and in part a modeled one; it is offered as a compounding reinforcement of the homeostatic valley over the soil's maturation and life, not as the proximate engine of the transition, which remains the proximity economy described above.
Multiplicative rather than additive. The proximity terms are not independent perks to be summed. Density enables specialization; specialization lowers per-capita cost; lower cost supports higher density and tighter coupling. The terms feed one another, which is what produces a threshold rather than a smooth ramp. This is the flywheel, and it turns on ecological time; beneath it the Black Queen clock turns across generations, so a state established by the fast loop is deepened by the slow one, and the system is reinforced on two timescales at once. Section 4 shows that such positive feedback is precisely the formal requirement for the regime shift the paper proposes.
The boundary of this section is that all of the above is established as mechanism, supported by theory and by laboratory and synthetic-community experiments. What it does not by itself demonstrate is that a real soil crosses into a discrete, higher-order community state. It shows that every gear the flywheel requires is real.
4. Crossing the threshold: critical transitions and alternative stable states
The intuitive notion of an ecological "escape velocity," a hump the community must clear before a higher-order organization becomes self-sustaining, has a rigorous and long-standing form in ecology: the critical transition across a tipping point into an alternative stable state.
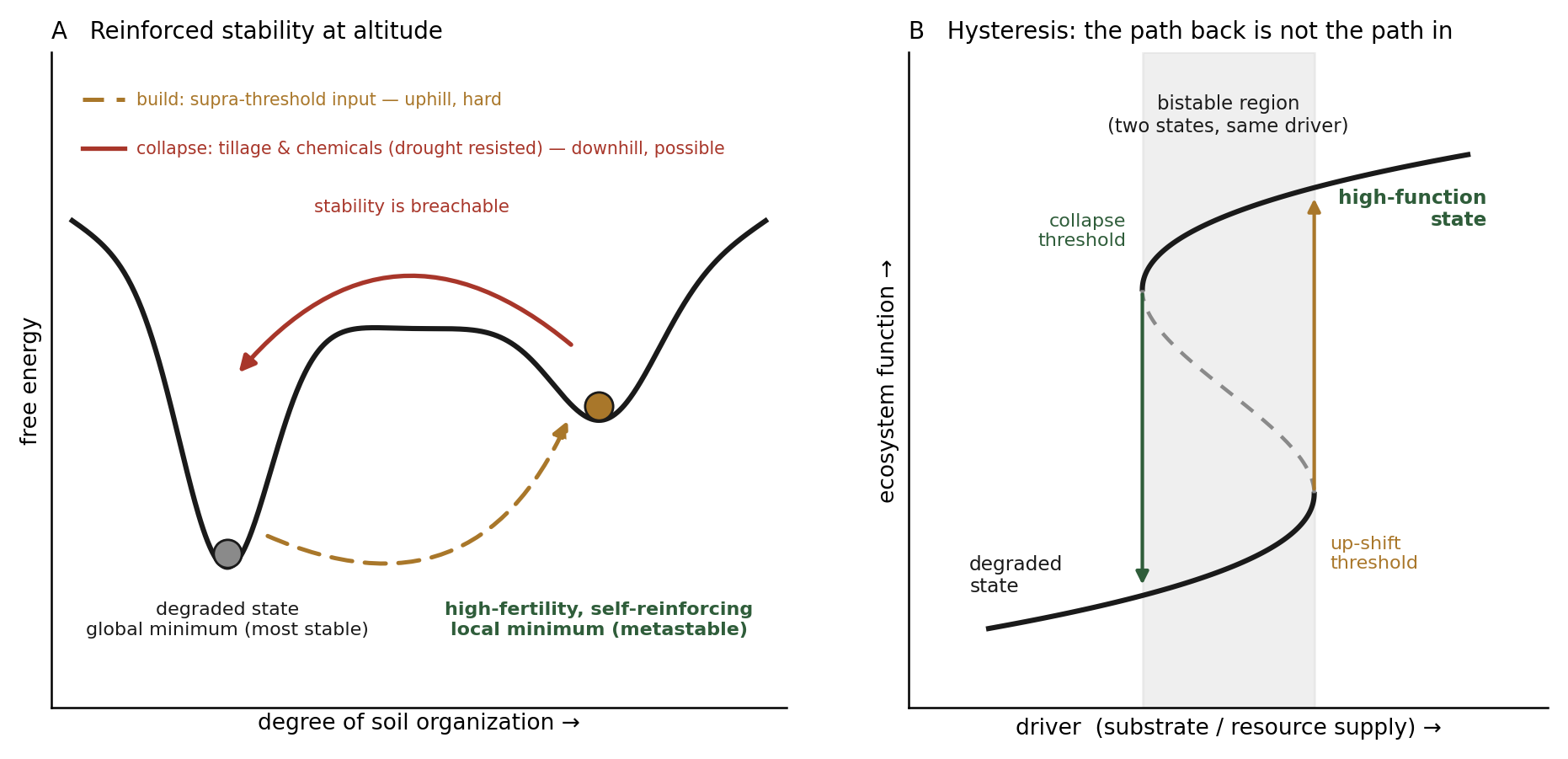
The theory holds that an ecosystem can persist in two or more stable states under identical external conditions; when a disturbance pushes it past a critical threshold, the response is nonlinear and disproportionately rapid, and the system shifts into an alternative state (Holling, 1973; May, 1977; Scheffer et al., 2001; Beisner et al., 2003). The "remains assembled" half of the intuition corresponds to hysteresis: once a tipping point is crossed, restoring the driver to its former value does not restore the former state, so a small change in conditions can produce an enduring change to a state with different characteristics. The familiar ball-and-cup image is apt (Figure 2). An "escape velocity" is the push required to move the ball over a ridge into a neighboring valley, not a runaway to infinity; once in the new valley, perturbations roll back toward it, which is the rigorous form of the homeostasis the hypothesis emphasizes.
Two features make this framework more than an analogy for the present argument.
First, positive feedback is a necessary but not sufficient condition for the emergence of alternative stable states at the community scale, since whether a positive feedback opens a second basin depends on its form and strength and not merely on its presence (Kéfi, Holmgren and Scheffer, 2016). The efficiency economy of Section 3 and the multifunctional surface of Section 2 are therefore not separate conveniences; in this framework they are the self-reinforcing loops without which a second basin cannot exist.
Second, a growing body of evidence locates the relevant transitions in microbial communities specifically, and increasingly in soil, not only in lakes and forests. Energy-landscape analysis of more than fifteen hundred agroecosystem soil samples finds that prokaryotic and fungal community structure resolves into several alternative stable states separated by inferred tipping points, with the states differing in their association with crop disease (Fujita et al., 2025). In controlled microbial communities the transitions have been driven and observed directly: a transient perturbation, including a temporary shift in pH, can induce a lasting switch between alternative stable states that persists after the perturbation is removed (Amor, Ratzke and Gore, 2020). Soil functional transitions show the same signature under moisture stress, where mild perturbation allows return to the reference state while intense perturbation pushes the community out of its stability basin into an alternative functional state that persists after the perturbation has ceased (Todman et al., 2018). That persistence after the cause is removed is the field-scale form of the self-stabilizing regime the hypothesis posits. The soil demonstrations to date indicate that such states exist and that transitions between them occur; experimentally tracing a full hysteresis loop in soil, rather than inferring the tipping points, remains the harder and still-open measurement.
The most directly relevant home for the argument is restoration ecology. For degraded systems with alternative stable states, the recovery trajectory is not the mirror image of the collapse trajectory, and feedbacks make restoration difficult; degraded systems become trapped in their state, so that reversing the original driver is insufficient to restore function. Stated in the field's own terms, this is the present argument: a depleted soil is not merely low on inputs but is held in a low-fertility basin by its own feedbacks, and cannot be coaxed out by incremental amendment. The rigorous justification for comprehensive, simultaneous intervention (carbon lattice, complete minerals, and inoculum delivered together as one supra-threshold input) is exactly this: half-measures relax back, whereas a threshold-crossing input can reorganize the system into the higher basin where it then holds. The most exact way to state what that intervention must be, kept as an analogy and not as evidence, is that an ecological community of this kind behaves like an engine rather than a pile of ingredients. Its parts, the durable carbon habitat, the complete mineral charge, and the living inoculum, must reach a minimum working complexity together before the self-reinforcing feedback will run at all, just as an internal-combustion engine below a minimum of compression, fuel, air, and ignition acting in concert does not run weakly but does not run at all, and only above that minimum does the flywheel carry itself. Incremental amendment is the engine cranked with no fuel in the line; the supra-threshold, all-at-once input is what allows it to catch. The same image, applied to the field record for biochar, returns in Section 8.
The synthesis that unifies the paper is that the carbon lattice is the durable abiotic anchor of the stabilizing feedback. Alternative stable states are typically driven by biotic-abiotic feedbacks. The lattice persists (Section 7), and while it persists it holds the water, nutrients, signals, and habitat (Section 2) that sustain the dense, interdependent, efficient community (Section 3), which in turn maintains the aggregation and organo-mineral integration that keep the system in the high-fertility valley. The roles here are distinct and should not be merged: the recalcitrant carbon is the durable anchor whose persistence keeps the stabilizing feedback supplied and therefore keeps the basin from eroding over time, but the basin itself, and its hysteresis, are produced by the self-reinforcing biology the substrate supports, not by the inert carbon. The carbon makes the state durable; the living feedback makes it an attractor.
These are two kinds of persistence, and conflating them obscures the claim. The carbon's durability is passive and kinetic, in the way an allotrope is: diamond sits above graphite, carbon's stable ground state at surface conditions, yet persists indefinitely because reverting would mean dismantling an ordered covalent network, a barrier so high the rate is negligible. Pyrogenic carbon is locked the same way, its recalcitrance rising with the order and condensation of its aromatic domains, which are turbostratic, nearer disordered graphite than cubic diamond, so the principle is borrowed and not the literal form. But a diamond does nothing to stay a diamond; it is trapped, not maintained, whereas the high-fertility function is active persistence, held by a living community that constructs the niche favoring it and is sustained only while energy flows through it. Terra preta couples the two: a diamond-like scaffold that persists passively and gives the engine a durable home, housing a flame-like community that supplies the fertility the scaffold alone cannot. This layering is why one soil shows both kinds of endurance at once, persisting for centuries and resisting drought through the inert scaffold, yet falling to tillage and biocides, which dismantle the living community rather than the carbon.
The persistence of terra preta for centuries after abandonment is consistent with this picture, but it is not by itself evidence of a living attractor, because recalcitrant carbon and passive cation exchange would endure for centuries with no self-reinforcing biology at all; endurance is therefore over-determined. What would distinguish a genuine self-stabilizing biological state from a durable but passive nutrient store is not that the soil lasts but how it lasts: a passive store is drawn down as harvests export nutrients, whereas a self-replenishing community would maintain fertility under continued cropping. That behavioral signature, persistence under export rather than persistence as such, is the discriminating test taken up in Sections 8 and 10.
Three boundaries must be stated honestly. An abrupt response alone does not prove bistability, because abrupt responses can also follow abrupt changes in external conditions; demonstrating an alternative stable state requires showing hysteresis, two states under the same conditions. The documented soil transitions are predominantly downward, into degraded states that are hard to restore; the deliberate up-transition into a high-fertility self-stabilizing basin is strongly plausible by the symmetry of the theory and is supported by the existence of dark earths as a built high-fertility endpoint, but it has not been demonstrated as a controlled experiment. And a real feedback is necessary but not sufficient; the loops must be strong enough to fold the system's response curve and open a second basin, and showing that they are that strong is the work that remains.
5. The fungal layer: integration, mineral mobilization, reciprocal exchange, and immune priming
Among the components of the proposed system the fungal network has the widest reach, and the right approach is to state its role at full strength and then to state, with equal clarity, where the evidence for that role ends. The functions resolve into five of differing evidentiary weight: the network integrates the system, mobilizes its minerals, runs a reciprocal market with its plant partners, primes plant immunity, and carries signals. The first four are well supported. The fifth is supported in part and carries one widely publicized overstatement that must be set aside.
Integrate the system (well supported). The fungal network is the closest thing the soil has to an integrating organ. Mycorrhizal and saprotrophic fungi govern large parts of the soil economy at once: they regulate nutrient cycling, shape microbial diversity, suppress soil-borne pathogens, and bind mineral particles and organic matter into the aggregates that give soil its structure, while their necromass and the carbon they receive from plants enter the stable organic-matter pool. The integrating role is measurable at the level of the whole system rather than merely inferred. In manipulative experiments the below-ground diversity of arbuscular mycorrhizal fungi was shown to determine plant diversity, ecosystem variability, and productivity, with the diversity, stability, and yield of the entire plant community shifting as the fungal community was altered (van der Heijden et al., 1998). This is the strongest sense in which the network organizes the system: a change in the fungal layer reorganizes the output of the whole. That organizing role is an emergent ecological function distributed through every layer of the soil, the connective and conductive tissue of the system, and it is best described that way rather than as a central controller sitting atop a hierarchy. One calibration on the carbon role specifically: it is dual, since fungal necromass stabilizes organic matter while mycorrhizal activity can also accelerate turnover, so the defensible statement is that fungi govern the carbon balance, not that they only store carbon.
Mobilize and shuttle minerals (well supported). The network is also the plant's principal organ for acquiring the limiting mineral elements. Hyphae extend the effective reach of the root system by orders of magnitude and acquire phosphorus, nitrogen, water, and trace elements that the root alone cannot reach, including elements released from mineral surfaces by fungal activity (Smith and Read, 2008). In a soil built on a durable, high-exchange carbon habitat, this acquisition network is the system's means of drawing mineral stock into biological circulation, and it is the same reach that the companion framework develops as a downward, in-place mining of the mineral substrate.
Run a reciprocal market with plant partners (well supported). The exchange between plant and fungus is not a passive leak of resources but a regulated, reciprocal trade. Plants can detect, discriminate, and reward the fungal partners that supply the most phosphorus by allocating them more carbon, and the fungi reciprocally direct more nutrient transfer to the roots that supply the most carbon, so that cooperation is stabilized and defection is penalized on both sides (Kiers et al., 2011). This is a biological market with partner choice and sanctions, and it is the rigorous, evidence-based content of the intuition that the system negotiates: discrimination and reciprocal reward, not deliberation. The calibration is that the reward mechanism is real but context-dependent, varying with the nutrient demand of the host and the supply on offer rather than operating as a fixed rule.
Immune-mobilize (well supported, and a literal immune function). Mycorrhiza-induced resistance (MIR) is a named, replicated phenomenon in which colonization by arbuscular mycorrhizal fungi produces a mild activation of the plant immune system, locally and systemically, leaving the plant "primed" so that defenses activate faster and more strongly when an attacker arrives, centered on the jasmonate-regulated defense pathway (Pozo and Azcón-Aguilar, 2007; Jung et al., 2012). MIR has been demonstrated across a broad range of pathogens and herbivores in many crops. The honest calibration is that the effect is context-dependent; under high-nutrient or high-light conditions the priming can be reduced or reversed, so the defensible phrasing is that fungi prime plant immunity under most conditions (Dejana et al., 2022).
Signal and communicate (graded, with one overstatement to set aside). The communicative claims must be separated by strength. The plant-fungus molecular dialogue is settled science: under phosphorus stress the plant exudes strigolactones that promote fungal germination and hyphal branching (Akiyama et al., 2005), and the fungus replies with lipo-chitooligosaccharide signals recognized by plant receptors that activate the common symbiosis pathway (Maillet et al., 2011). Beyond that point the evidence weakens and must be labeled honestly. Individual experiments report that plants joined by a common mycorrhizal network can transmit defense cues, with connected receivers showing raised disease resistance or emitting defensive volatiles when a donor is attacked (Song et al., 2010; Babikova et al., 2013); these results are suggestive, but they are also among the single-study network-signaling findings that the systematic review noted below found to be over-cited and not yet robustly established, and they are presented here as contested rather than secure. Electrical activity resembling action potentials has been recorded propagating in hyphae (Adamatzky, 2022); this is a real phenomenon, but its interpretation as a language is unsupported speculation and is not evidence of cognition, and it is treated here as an open frontier at most. The romantic synthesis of these threads, that forests are densely mapped fungal internets through which mature trees consciously share resources and warnings with their kin, was formally challenged by a systematic review that found the claims of widespread networks, of reliable resource transfer that benefits recipients, and of preferential parent-to-offspring provisioning to be insufficiently supported or, in the case of the last, without peer-reviewed evidence (Karst, Jones, and Hoeksema, 2023). That narrative is set aside, and the molecular dialogue, the immune priming, and a cautiously stated and contested interplant signaling are retained.
The point that ties this layer to the rest of the paper is that these fungal functions scale with mycelial density: interplant transfer through the network has been reported to respond to the density of the mycelial network. If the carbon lattice supports a denser fungal network (Sections 2 and 3), then by this relationship the integrating, acquisitive, exchange, immune, and signaling functions all intensify with that density. The fungal layer is not a separate claim; it is one more set of functions riding the same surface-area and density flywheel, and the "real estate" thesis predicts the very variable observed to control them.
Stated plainly, then, the evidence supports a fungal network that integrates the system, acquires and mobilizes its minerals, trades reciprocally with its plant partners, and primes their immunity, with all of these functions intensifying as the network densifies. That is a commanding organizing role, and the paper rests on it. What the evidence does not support, and what this paper does not claim, is that the network is intelligent, that it deliberates, or that it consciously manages the system from above. The organization it provides is the emergent product of distributed function and reciprocal exchange, the integrating tissue of the soil rather than its mind, and stating the role at exactly that strength is what keeps it defensible.
6. Convergent mechanisms: five fields, one architecture
The strongest support this synthesis can offer is not any individual citation but a pattern of convergence. Five disciplines that do not generally cite one another each describe a component of the same architecture: dense, structured, resource-replete communities crossing feedback-driven thresholds into self-stabilizing, high-function states. When independent fields, studying different systems, each recover a piece of one architecture, that is good evidence that the components are real and that the architecture is assembled from established parts. It is not, by itself, evidence that those parts assemble into the integrated outcome in a soil, which is the step Section 10 isolates as the open experiment. What the convergence establishes is that the hypothesis is built from accomplished science rather than speculation, and that is what makes it worth the test.
Disease-suppressive soils (microbiology). A suppressive soil is one in which a soil-borne disease fails to develop despite the presence of a virulent pathogen, a susceptible host, and favorable conditions, because of the microbial community; the phenomenon has been studied for decades (Weller et al., 2002) and has been framed as a soil immune response (Raaijmakers and Mazzola, 2016). It comes in two forms whose properties map onto this paper's claims: general suppression, driven by total microbial biomass, an inherent and non-transferable soil property (the density claim, expressed as community-level immunity), and specific suppression, attributable to particular microbes and transferable by mixing a small fraction of suppressive soil into a conducive one, and eliminable by heat (a near-demonstration of state transfer by inoculation, which is the intervention model). The calibration is that general suppression is correlative rather than a switchable state, and that the occurrence of suppressive soils is often difficult to predict.
Pore-scale coexistence (soil physics). Microfluidic experiments in soil-aggregate-mimicking pore networks show two bacterial species, an obligate aerobe and a facultative anaerobe, segregating into preferred regions along opposing carbon and oxygen gradients and coexisting persistently in structured space in a way that is not possible in well-mixed culture (Borer, Tecon and Or, 2018). This is the skyscraper-versus-off-grid intuition demonstrated directly: structured space sustains coexistence that homogeneous mixing destroys. Consistent with that result, soil is understood to organize into microbial hotspots of locally intense activity, concentrated at sites such as the rhizosphere, the detritusphere, biopores, and aggregate surfaces, with the scale of those hotspots set by the diffusion of substrates and signals through the pore structure (Kuzyakov and Blagodatskaya, 2015), which fuses the surface argument of Section 2 with the economy of Section 3.
Priority effects and community assembly (community ecology). The order and timing of species arrival cause historical contingency in community structure and can produce alternative stable states through niche preemption and modification (Fukami, 2015). Which basin a community lands in can therefore depend on who arrives first, which sharpens the intervention model: it may not be enough to add resources and biota; the desired community may need to be established first and fast so that it preempts the niches before the degraded community reasserts itself. The honest crack is the live debate over how deterministic versus stochastic assembly is, and the fact that some perturbed systems recover toward their reference state, so a new state must be genuinely self-stabilizing rather than transiently different.
Plant-soil feedback (agroecology). Plants condition soil and soil conditions the next plants, a feedback loop now being developed deliberately as a tool. A proof-of-concept showed that steering the soil microbiome through plant-soil feedback can induce above-ground resistance to an insect pest in a crop (Pineda et al., 2020), and a recent synthesis argues for applying plant-soil-feedback principles to restore soil function in agriculture (Wang et al., 2025). This connects suppressive soils, priority effects, and the present thesis: conditioning produces a suppressive or immune community that can be steered by inoculation. The crack is that feedback can be negative as well as positive, and that how to steer microbiomes toward their beneficial functions remains largely unknown.
Comparative dark earths (anthropogenic soils). Terra preta is not unique to Amazonia, and, critically, the method is not entirely lost. West African communities produce carbon-rich, high-fertility African Dark Earths today, analogous to Amazonian terra preta, by accumulating ash, char, bones, and organic residues in kitchen gardens and middens (Solomon et al., 2016). These soils hold substantially more organic and pyrogenic carbon, more available phosphorus, and higher cation exchange capacity than adjacent soils. The honest crack is the persistent intentionality debate: it remains likely that Amazonian terra preta was not deliberately engineered for fertility at large scale, so the intentional, engineered-at-scale version is the thing still to be demonstrated, which is again the open experiment.
The unifying observation is the convergence itself. Microbial ecology, soil physics, community ecology, ecosystem theory, and restoration ecology each, in their own systems, describe a piece of one density-and-feedback-driven transition into a self-stabilizing high-function state. That convergence is what makes the hypothesis credible and worth testing, independent of any single source; what it does not do is stand in for the integrated demonstration, which remains the open experiment of Section 10.
7. The carbon question: persistence, decomposition, and the limits of "permanent"
The mechanism in Sections 4 and 6 depends on the carbon lattice persisting long enough to anchor the stabilizing feedback. This raises two related questions: how stable the lattice actually is, and whether anything in soil is truly permanent.
The modern view of soil organic matter persistence directly supports the framing used here. The field has largely abandoned the idea that certain molecules are intrinsically un-decomposable; persistence is now understood as an ecosystem property arising from environmental and biological context (physical inaccessibility, mineral association, the local absence of the right organisms or enzymes, redox, and moisture) rather than as a fixed molecular property (Schmidt et al., 2011; Lehmann and Kleber, 2015). Pyrogenic carbon fits this picture as an unusually durable but still finite material. Isotopic and incubation studies partition biochar carbon into a small labile pool that mineralizes within months and a dominant recalcitrant pool with a mean residence time of centuries, with relatively little carbon released over multi-year tracing studies (Kuzyakov et al., 2009; Wang et al., 2016). The condensed aromatic framework is that durable backbone.
There are three distinct fates for the lattice, and only one resembles simple reordering. The labile fraction is genuinely oxidized to carbon dioxide and lost (true mineralization, larger in lower-temperature chars). The surface is also oxidized, but productively, forming the oxygen-bearing functional groups that build cation exchange capacity over time. The condensed core largely persists, slowly, for centuries. For the purpose of building a high-fertility soil, the early labile loss is modest and much of the breakdown is itself the maturation process: fragmentation exposes fresh surface, surface oxidation builds exchange capacity, the labile carbon feeds the early microbial bloom, and condensed fragments integrate with minerals into organo-mineral complexes. For carbon-permanence accounting, by contrast, the cost of a lower-temperature char is real, because permanence rewards the most recalcitrant material. One context-specific note is that pyrogenic carbon may turn over faster in tropical Ferralsols than global averages suggest, which would imply faster maturation but shorter permanence in such soils.
A further part of the lattice's contribution to durable carbon is indirect. A substantial share of the organic carbon stored in the anthropogenic dark earths is non-pyrogenic, and recent carbon-partitioning work finds that their carbon enrichment cannot be attributed to pyrogenic material alone (Solomon et al., 2016; Slocum et al., 2026). Consistent with the ecosystem view of persistence, the durable carbon and the mineral surfaces it helps organize plausibly stabilize co-located humified organic matter, extending its residence time beyond what the molecules themselves would have (Schmidt et al., 2011; Lehmann and Kleber, 2015). The lattice's durable-carbon role is therefore partly its own recalcitrance and partly the protection it lends the non-pyrogenic pool around it, which also means that a complete soil-building input includes the humifiable organic matter that pool is built from, not the lattice alone.
On the broader proposition that nothing is permanent given enough time and fuel, the honest position is that this is more right than wrong, but the absolute version overshoots, and the place it breaks is instructive. Time erodes kinetic barriers, so slow reactions eventually proceed; time does nothing to energetic barriers. A transformation that is thermodynamically uphill at ambient conditions will not proceed microbially regardless of elapsed time or unrelated available energy, unless it is coupled to a sufficiently exergonic partner, and there is a minimum quantum of free energy a cell can conserve, on the order of twenty kilojoules per mole, below which no organism conserves energy and therefore no enzyme evolves (Schink, 1997). This is why "given enough fuel" misframes the problem: the organism must conserve energy from the target transformation itself or its coupled partner, not from arbitrary calories nearby. Fully oxidized minerals offer nothing to oxidize for energy, and the carbon-fluorine bond of per- and polyfluoroalkyl substances is a clean modern example of a material that resists biodegradation for want of an energetic handle. The most condensed carbon, including the aromatic core of the lattice, presents few accessible edge or defect sites for attack, which is precisely why that fraction is the most persistent.
The useful synthesis is the one the field itself adopts: "permanent versus breakable" is the wrong binary, and "rate of decomposition under these specific conditions" is the right one. Under that framing, the philosophical point holds (almost nothing is infinitely permanent) while the absolute claim does not (a few things are effectively inert for want of energy or a handle, and thermodynamics is a wall that time does not sand down). The practical payoff is that soil-carbon residence times span roughly twelve orders of magnitude, and that spread is the entire value proposition: a lattice that persists for centuries while slowly feeding the system is what makes the mechanism work agronomically, and it is exactly the caveat that must attach to any permanence-based carbon claim. The craft is not making the lattice permanent or breaking it, but tuning its rate.
One implication of this durability belongs to the argument that follows and is best stated plainly here. Precisely because the lattice and its passive exchange capacity endure on their own for centuries, the mere endurance of a dark earth cannot serve as evidence that a living, self-reinforcing community is holding it in place, since the carbon explains the longevity with no help from biology. The biological claim of this paper therefore does not rest on persistence as such. It rests on how a soil persists, the distinction developed in Sections 8 and 10.
8. Competing explanations: the abiotic account and the biochar record
A hypothesis of this kind earns its place only by facing the most economical rival explanation and the mixed empirical record honestly, rather than by collecting confirmations. For terra preta and for biochar there is a strong rival and a genuinely uneven field record, and both are taken up here directly, because both, read carefully, narrow the claim rather than defeat it.
The rival explanation is abiotic, and it is the parsimonious null. On this account the fertility and the persistence of dark earth are products of chemistry rather than of any change in biological organization: large stocks of phosphorus and calcium from bone, ash, and waste; a high cation exchange capacity that holds nutrient cations; a raised and buffered pH; and a recalcitrant carbon skeleton that endures for centuries. This reading is reinforced by how the soils are now understood to have formed: archaeological and ethnographic work traces Amazonian dark earth to the sustained deposition of nutrient-rich residues — fish and animal bone, ash, charcoal, and domestic refuse — accumulated in and around settlements over generations, whether as deliberate soil improvement or as a byproduct of habitation (M. J. Schmidt et al., 2023). That formation history is the strongest form of the abiotic account: a centuries-long nutrient subsidy, not a single engineered event, and the present claim that the state can be founded quickly and then largely sustains itself must be measured against it rather than assumed past it. The African dark earth data are consistent with this reading, since those soils carry several-fold to many-fold more available phosphorus, a higher exchange capacity, and a more favorable pH than their neighbors (Solomon et al., 2016), and the biochar meta-analytic record attributes a large part of biochar's yield effect to a liming effect rather than to anything subtler (Jeffery et al., 2017). The honest position is that this account is real and that it may explain a large share, perhaps the majority, of what makes dark earth fertile and durable. A responsible version of the present hypothesis does not deny the abiotic contribution; it states precisely what it adds to it.
What it adds is an answer to two things the abiotic account does not by itself explain. The first is self-maintenance under export. A nutrient bank with high exchange capacity is still a stock, and a stock is drawn down as harvests carry nutrients off the field, so passive chemistry predicts slow depletion rather than the maintenance of fertility under continued use. A living community that fixes nitrogen, solubilizes phosphorus, and rebuilds organic matter is the kind of system that could keep a soil replenished rather than merely well-stocked, and self-replenishment, not storage, is the property that would distinguish a soil in a self-sustaining state from a rich but finite bank. The second is the shape of the response. Additive chemistry predicts that benefit scales smoothly with input and reverses when the input is withdrawn, whereas the regime-shift claim predicts a threshold, a disproportionate response once it is crossed, and persistence afterward. Terra preta is also observed to host distinct and persistent microbial communities, different from those of adjacent soils (Kim et al., 2007), which is consistent with a distinct biological state, though on its own it cannot show that the biology is a cause rather than a consequence of the chemistry. The two are separated not by assertion but by measurement: whether the high-function state persists under continued export, whether it shows threshold and hysteresis behavior rather than smooth reversibility, and whether a phosphorus fractionation time course finds phosphorus held in biologically re-releasable pools rather than passively adsorbed or lost to occlusion. These are the experiments of Section 10, and they are designed to test the biological claim against the abiotic null rather than to assume it.
The field record for biochar must be read with the same honesty, because at first glance it looks unfavorable to any strong claim. Across meta-analyses the average yield effect is modest, on the order of ten to thirteen percent, and highly variable, with many null and some negative results (Jeffery et al., 2011; Schmidt et al., 2021). Part of that variability is publication-sensitive, and at least one analysis found a headline benefit reverse its sign after correction for publication bias. None of this is set aside here. But the same record carries a pattern that is the opposite of discouraging for this hypothesis: the benefit is concentrated in acidic, sandy, low-exchange, weathered soils in tropical climates, with large gains where the initial pH is low and little or no effect in neutral, fertile, temperate soils (Jeffery et al., 2017; Schmidt et al., 2021). That is not random scatter. It is the dark-earth setting itself, the soils in which the mechanism proposed here should matter most and in which ordinary ground has the least to begin with.
The remaining heterogeneity is, to a large degree, an artifact of how biochar has been tested, and this is where the preparation argument of Section 2 becomes decisive rather than rhetorical. Most trials apply fresh, uncharged, uninoculated char and measure the result over one or two seasons. By the sink-to-source argument that is precisely the artifact, and precisely the interval, in which char should perform worst, because an uncharged lattice first draws nutrients and water out of solution before it returns them. The prediction is borne out directly: in a controlled comparison, untreated biochar reduced plant biomass to sixty percent of the unamended control, while the same biochar, co-composted and thereby nutrient-charged before application, raised biomass by up to three hundred percent in the same poor soil, an effect traced to captured nitrate and phosphate held against leaching and released slowly (Kammann et al., 2015; Joseph et al., 2018). Fresh char and charged char are, functionally, two different materials, and a literature that has largely tested the first cannot be read as a verdict on the second.
The same engine image introduced in Section 4 is the most exact way to read this record. Most biochar trials are not weak tests of the assembled system but tests of a disassembled one: a bare lattice, or a lattice plus a pH correction, run for a season or two, is an engine turned over with no fuel in the line and no spark, and its failure to start is not a verdict on the engine. The claim is not that assembly guarantees ignition, which is precisely what the experiment in Section 10 is built to find out, but that a record compiled almost entirely from incomplete assemblies cannot adjudicate a hypothesis about the complete one.
This argument must not be allowed to become unfalsifiable, and it is not. It does not claim that every null result reflects improper preparation; some soils are already fertile and buffered and have little to gain, which the meta-analyses show plainly. The claim is narrower and testable: that charged, aged, and inoculated char will outperform fresh char, that the gap will widen over years rather than seasons, and that the comparison which would settle the matter, fresh against charged against charged-and-inoculated, tracked over time and against an explicit liming control, has rarely been run. That same comparison is what separates the biological claim from the abiotic one, which is why the experiment in Section 10 carries the weight of the paper. The mixed record is not evidence against the hypothesis. It is evidence that the decisive experiment has not yet been done.
9. Emergent efficiency: resource economy as a consequence of the mechanism
The mechanism advanced above predicts more than persistence; it predicts a distinctive resource-use efficiency, and predicts it as a single emergent property rather than a set of separately engineered benefits. A community that has crossed into the self-reinforcing basin (Section 4) is, by construction, one in which the limiting resources are retained, recycled, and mobilized rather than lost — that retention is what holds the basin in place. Efficiency is therefore not an add-on to the mechanism but its expected signature, and the dimensions below are facets of the same organization. Each is stated as a mechanistic prediction whose magnitude is the open question the experiment that follows is designed to quantify; none is asserted as demonstrated.
Water economy follows directly from the retention functions of Section 2. The lattice and the biological aggregation it supports raise water-holding capacity and intercept the leaching flux, reducing evaporative and drainage losses and buffering the community against dry spells; the floor on this efficiency is set by the transpiration the crop must run to fix carbon, not by the soil, so the predicted gain is largest in coarse, low-clay soils with the least retention to begin with. Nutrient economy follows from the same surface and the community it houses: microbial immobilization and fungal transfer recycle nutrients internally, while the charged lattice holds mineral nitrogen and other ions against leaching and denitrification (Kammann et al., 2015). The prediction is a reduced external replacement rate, not a closed loop — what the harvest exports must still be supplied — and the distinction between a tightly recycling system and a passive store is precisely the persistence-under-export signature developed in Section 8.
Two further efficiencies are properly biological. The system can supply part of its own nitrogen through diazotrophic fixation in association with the plant–fungal network, an input limited not by atmospheric supply but by the delivery of N₂ to the fixers and the protection of nitrogenase from oxygen — conditions a well-aggregated, biologically ventilated topsoil is structured to meet. And metabolic efficiency rises as complete mineral nutrition restores the micronutrient cofactors of the photosynthetic and respiratory machinery, moving the limiting efficiencies toward their biological ceilings; the gain over depleted ground is large precisely because depleted ground operates far below those ceilings. None of these efficiencies creates resources. They convert and conserve them — the system runs efficiently on the fuel it is supplied, and the measure of how efficiently is the falling maintenance-input curve of the experiment that follows.
10. From synthesis to demonstration: the decisive experiment
The preceding sections show that every component the proposed mechanism requires is independently established, and that five disciplines converge on its architecture. What they do not show is the integrated outcome: that a real soil can be driven across a threshold into a self-stabilizing, terra-preta-like state. The synthesis is the groundwork; the demonstration would be the discovery. The design that follows is built to discriminate the specific claim of this paper from the strongest competing account, because it is not enough to show that a matured soil holds high function when inputs are withdrawn. A passive carbon-and-exchange bank, the abiotic account conceded in Section 8, predicts exactly that: a soil amended with recalcitrant carbon and base cations holds its gains for a long time simply because the carbon does not decay and the exchange sites do not empty. The living-attractor claim makes a stronger and different prediction, and the experiment must isolate it.
The discriminating variable is not withdrawal of inputs but continued export. A passive bank, however large, is drawn down by repeated cropping: each harvest removes nutrients, and with no resupply the store declines on a schedule set by its size. A soil whose fertility is maintained by a self-replenishing community predicts the opposite signature, high function sustained across repeated harvest cycles, because the community continues to weather minerals, retain mobile nutrients, and rebuild the pools faster than a passive store would allow. The decisive contrast is therefore performance under sustained cropping with external inputs withheld, run long enough for a passive bank to draw down measurably. Persistence under export, not persistence under rest, is the signature of a living attractor, and it is also the operational meaning of the paper's claim that replication holds.
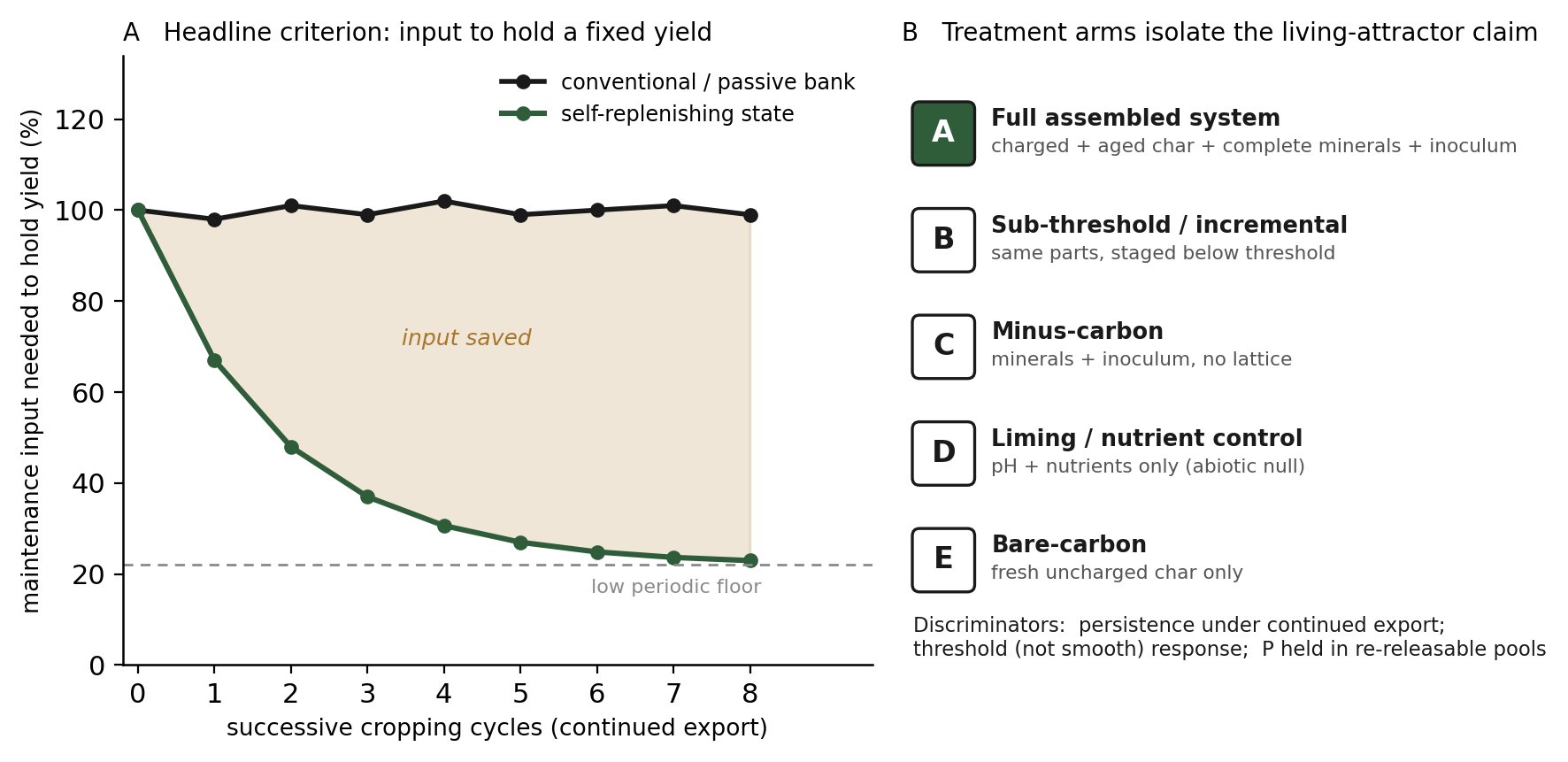
There is a second and more legible way to read the same signature, run as a complementary protocol alongside the persistence arms, and it is the one most useful as a headline success criterion. Rather than withholding inputs entirely and watching for persistence, a parallel set of plots, held at a fixed yield by titrated maintenance, can track how much input that constancy requires across successive cropping cycles. A passive bank, and equally a conventionally fertilized control, requires a roughly constant input to hold yield, because what is exported must be replaced in full each cycle. A soil that has crossed into a self-replenishing state predicts a different curve: the maintenance input needed to hold the same yield falls over successive cycles toward a low, periodic floor (Figure 3), as the installed exchange capacity and the living community take over the work of retaining and recycling nutrients, leaving only the most leachable and exported elements, chiefly potassium and then phosphorus, to be topped off. A declining maintenance input at constant yield is the affirmative signature of a founding charge that then largely sustains itself; a flat or rising input is its refutation. This curve is the cleanest single result the program can place before a prospective research partner, because it states the whole claim, that fertility can be founded once and then mostly holds itself, in a form that is directly measured rather than inferred.
The design is a single common-garden experiment on a degraded, low-carbon, low-exchange soil of the kind in which the mechanism is most relevant, with a real terra preta sample as a reference endpoint where available. Every arm receives the same total mineral loading and is brought to the same pH and base saturation, so that the carbon lattice and the biology, not nutrients or acidity, are the variables under test. The arms are: (A) the full treatment, conditioned carbon matured to high exchange capacity and mineral-loaded, plus a defined and locally augmented inoculum, at a supra-threshold dose; (B) the same full package at a sub-threshold dose; (C) a minus-carbon arm receiving the identical minerals and inoculum with no carbon lattice, optionally with an inert high-surface filler to separate physical structure from pyrogenic carbon; (D) a correction control brought to the same pH, base saturation, and nutrient status by liming and soluble fertilizer alone, without the lattice and without the augmented inoculum, representing conventional agronomic practice; and (E) a bare-carbon arm receiving fresh, unconditioned char without mineral loading or inoculum. An untreated degraded-soil baseline completes the set.
The readout is the whole terra preta syndrome measured together, not yield alone, because the central proposition is that the biological properties emerge as a set from one substrate rather than being installed one at a time. Across the cropping cycles each arm is scored for crop performance, microbial density and community diversity and network complexity, disease suppression and markers of induced plant immunity, aggregate stability and tilth, water-holding capacity, and nutrient retention. Co-emergence of the suite in the full arm, appearing over time from substrate and inoculum without each function being engineered separately, is the positive signature of emergence; if the properties appear only when installed individually, that part of the claim fails.
The contrasts decide the question. If the full arm (A) sustains high function under continued export while the sub-threshold arm (B) relaxes toward baseline, the result is a history-dependent alternative stable state with hysteresis, two outcomes from the same endpoint conditions. If the full arm holds while the minus-carbon arm (C) fails to establish or fails to sustain the biological dividend, carbon is demonstrated to be the keystone variable rather than one contributor among equals. If the full arm holds under export while the correction control (D) decays as its soluble store is cropped away, the outcome is shown to be more than a pH-and-nutrient correction, the dimension on which the abiotic account was granted its force in Section 8. If the full arm outperforms the bare-carbon arm (E), the mineral loading and biological charging of the lattice are shown to matter, consistent with the contrast between fresh and co-composted char in the literature (Kammann et al., 2015). The physical dividend, by contrast, is expected in every carbon-bearing arm immediately; it is not what the experiment is testing.
The cross-disciplinary sweep also sharpens how to attempt the up-transition rather than only how to test it. Suppressive-soil transfer indicates that the target community can be moved as an inoculum (Section 6), and priority-effect theory indicates that timing is decisive: the desired community should be established early enough, and at sufficient density, to occupy the niches before the degraded community reasserts itself, which is the operational form of the founding-conditions bound stated in the introduction. That the endpoint can be built by hand is not in doubt, since high-fertility dark earth has been produced repeatedly by traditional practice in West Africa and Amazonia, an existence proof for the endpoint even if not for the controlled, instrumented, engineered route. What has not been done is the demonstration of that route with the internal controls above.
Two supporting measurements convert specific mechanistic claims into numbers within the same design. A sequential phosphorus fractionation run as a time course, resolving labile, iron- and aluminum-adsorbed, apatite-bound, and occluded pools, would show whether the system parks phosphorus in re-releasable pools or loses it into the occluded pool, and a leaching column would test the anti-leaching claim directly. Varying the weathered-clay fraction across otherwise identical full-treatment replicates would locate the point at which an oxide surface flips from a re-chargeable phosphorus buffer into a net sink that immobilizes both phosphorus and the microbial acids that would otherwise free it, the knife-edge that determines whether a phosphorus-fixing matrix is an asset or a liability in a biologically intense soil.
The design is falsifiable in each of its claims, which is its purpose. A full arm that decays under continued export at the rate of the correction control would refute the living-attractor claim and vindicate the passive-bank account. A minus-carbon arm that matches the full treatment would refute the keystone-variable claim. A supra-threshold arm (A) that does no better than the sub-threshold arm (B) would refute the minimum-complexity claim itself, collapsing the prediction back to the smooth, incremental dose-response that most amendment trials report and against which the threshold claim is deliberately staked. A correction control that reproduces the whole suite and holds it under cropping would show that liming and fertilizer suffice and that neither the lattice nor the biology does the work the paper assigns them. The one outcome the design does not permit is a free pass for a null result: because the controls are internal and the arms differ one factor at a time, a failure of the full arm cannot be set aside as poor char or weak inoculum without that same explanation being tested against the other carbon-bearing arms.
11. Conclusion
Terra preta is best understood not as a fertilized soil but as a soil whose biological community has settled into a different equilibrium, and the proposed mechanism for that equilibrium is a durable, high-surface-area carbon lattice acting as a multifunctional shared substrate. The lattice supplies habitat, exchange capacity, reservoirs, signal retention, and shelter through different pore classes and surface chemistries that ripen on different clocks; the resulting density and proximity switch on an efficiency economy governed by quorum sensing and a division of labor whose profitability hinges on proximity, with the Black Queen dynamic of adaptive gene loss compounding that economy across a longer evolutionary clock; the positive feedbacks of that economy are the formal prerequisite for an alternative stable state, a regime that a growing body of evidence indicates soil microbial communities can occupy; and the fungal network integrates the system, mobilizes its minerals, trades reciprocally with its hosts, and primes plant immunity, with these functions scaling as the mycelial network densifies. Five independent disciplines describe components of the same architecture, and that convergence is the strongest support presently available, though it motivates the hypothesis rather than demonstrating it.
The honest verdict is that the synthesis is strong, defensible, and testable, that its components are individually well established, and that the integrated outcome (an engineered, hysteretic up-transition into a self-stabilizing high-fertility state) is the open experiment that would distinguish a compelling hypothesis from a demonstrated discovery. The calibrated version of the claim is also the more powerful one, because the corrections that this synthesis incorporates (a tipping point rather than a runaway, an efficient route rather than a unique one, asynchronous maturation rather than instant function, measurable rather than conscious fungal signaling) are exactly what make the result survive scrutiny.
The strength of the case lies where it should: in the convergence of five independent fields on a single, testable architecture, and in the experiment that would convert that architecture from a coherent hypothesis into a demonstrated result. The applied and societal consequences of the mechanism, the deliberate building of such soils and the stakes of doing so, lie beyond the scope of this paper, which ends where the science does, at the threshold of that experiment.
Table 1. Evidentiary status of the principal claims advanced in this paper. Established and well-supported claims are stated plainly; partial claims are flagged; framings to be used sparingly or avoided are marked as such.
| Claim | Status | Principal support | What would strengthen it |
|---|---|---|---|
| Char adds large total surface area to low-surface-area soils | Established | Comparative surface-area data for chars vs. kaolinitic/sandy soils | Direct measurement of the specific char in use |
| Order-of-magnitude surface-area gain | Well supported, conditional | Holds for low clay baselines and high-area chars; several-fold otherwise | Stating it as conditional on soil and char properties |
| Habitable surface is macropore-limited; gas-adsorption area overstates it | Established | Accessible-fraction and pore-size colonization data | Pore-size distribution of the specific char |
| Surface, CEC, and habitat are partly independent design levers on different clocks | Well supported | Pyrolysis-temperature and aging chemistry; feedstock-morphology control of macropores | Time-course CEC and habitat measurements |
| Density-gated collective switching (quorum sensing) | Established | Miller and Bassler (2001); Waters and Bassler (2005) | Phrasing the threshold as "threshold-like," not a hard wall |
| Confinement lowers the density required (diffusion sensing) | Well supported | Redfield (2002) | Pore-scale confirmation in a soil-like matrix |
| The Black Queen dynamic compounds the proximity economy across an evolutionary clock, deepening the basin, and is not the proximate engine of the transition | Established as evolutionary theory; constructive form modeled | Morris, Lenski, and Zinser (2012); Takeuchi et al. (2024) | An evolutionary-time, partly modeled layer; demonstration in a soil community |
| Inoculated communities are already pervasively auxotrophic and cross-dependent, so the interdependent web is inherited rather than evolved in place | Established | D'Souza et al. (2014); D'Souza and Kost (2016) | None needed |
| Proximity flips division of labor from net cost to net gain | Well supported | Tsoi et al. (2018) | Direct soil-matrix test of the transport-penalty effect |
| Positive feedback is the prerequisite for an alternative stable state (necessary but not sufficient) | Established | Critical-transition theory (Holling 1973; May 1977; Scheffer et al. 2001); Kéfi, Holmgren and Scheffer (2016) | Showing the feedback is strong enough to fold the response curve |
| Soil microbial communities occupy alternative stable states; transitions show hysteresis | Well supported, growing | Fujita et al. (2025, soil energy landscapes); Todman et al. (2018, soil functional transitions); Amor, Ratzke and Gore (2020, experimental microbial transitions) | A full hysteresis loop traced experimentally in soil, not only inferred |
| Terra preta as an engineered, hysteretic up-transition | Frontier (open experiment) | Symmetry of the theory; existence of dark earths as a built endpoint | The hysteresis experiment in Section 10 |
| Fungal network integrates the system; its diversity determines plant diversity, stability, and productivity | Well supported | van der Heijden et al. (1998); soil fungal ecology reviews | Carbon role is balance, not storage only; the organizing role is emergent function, not control |
| Fungal network mobilizes and shuttles minerals to the plant (the principal acquisition organ) | Well supported | Smith and Read (2008) | None needed |
| Plant-fungus exchange is a reciprocal-reward market with partner choice and sanctions | Well supported | Kiers et al. (2011) | The "negotiation" is discrimination and reward, not deliberation; context-dependent |
| Mycorrhiza-induced resistance (fungi prime plant immunity) | Well supported | Pozo and Azcón-Aguilar (2007); Jung et al. (2012) | Noting context-dependence (Dejana et al. 2022) |
| Plant-fungus molecular dialogue | Established | Akiyama et al. (2005); Maillet et al. (2011) | None needed |
| Interplant defense signaling through fungal networks | Suggestive but contested | Song et al. (2010); Babikova et al. (2013), among the studies the systematic review cautions against | Present as contested, not secure |
| Fungal electrical activity in hyphae | Frontier; not evidence of cognition | Adamatzky (2022) | Real phenomenon; the "language" and intelligence reading is unsupported |
| Conscious, altruistic, forest-scale resource sharing ("wood-wide-web") | Avoid | Refuted as overstatement by Karst, Jones, and Hoeksema (2023) | Not applicable; drop the claim |
| The network's organizing role is emergent function and reciprocal exchange, not intelligence, deliberation, or conscious management | Calibration (limit) | Karst, Jones, and Hoeksema (2023) | State it as integrating tissue and function, never as cognition or a capstone-mind |
| Disease-suppressive soils as community-level immunity, transferable by inoculation | Well supported | Weller et al. (2002) | Predicting where suppression will occur |
| Dark earth is reproducible beyond Amazonia and made today | Well supported | Solomon et al. (2016); West African Dark Earths | Documenting an intentional engineered route |
| Pyrogenic carbon persists for centuries but is not permanent | Established | Schmidt et al. (2011); Wang et al. (2016); Kuzyakov et al. (2009) | None needed |
| Persistence alone is not evidence of a living attractor; endurance is over-determined by recalcitrant carbon and passive exchange capacity | Calibration | This synthesis; Schmidt et al. (2011) | The living-state signature is persistence under export, tested in Sections 8 and 10 |
| Durable carbon stabilizes co-located non-pyrogenic organic matter; dark-earth carbon enrichment is not exclusively pyrogenic | Well supported | Solomon et al. (2016); Slocum et al. (2026); SOM-persistence theory (Schmidt et al. 2011; Lehmann & Kleber 2015) | Direct demonstration that the lattice causes the co-located stabilization |
| Abiotic chemistry (phosphorus, calcium, exchange capacity, pH, recalcitrant carbon) explains much of dark-earth fertility and persistence | Leading alternative, conceded | Solomon et al. (2016); Jeffery et al. (2017) | The biological claim must be tested against this null, not assumed above it |
| Fresh, uncharged char can reduce yield, while charged or co-composted char improves it | Established | Kammann et al. (2015); Joseph et al. (2018) | None needed |
| Biochar's average field effect is modest and variable, and concentrated in acidic, sandy, weathered tropical soils | Established | Jeffery et al. (2011, 2017); Schmidt et al. (2021) | Liming and publication-bias confounds are real and acknowledged |
| The charging-and-maturation account explains much of the biochar literature's heterogeneity | Hypothesis (testable, not a blanket dismissal) | This synthesis; Kammann et al. (2015) | The fresh-versus-charged-versus-inoculated comparison, tracked over years against a liming control |
| A thermodynamic floor limits "anything decomposes given time and fuel" | Established | Schink (1997); PFAS recalcitrance | None needed |
| High-function state self-organizes under selection; its endpoint is shaped by founding conditions and local biota (priority effects) | Well supported | Community-assembly and priority-effect theory (Fukami, 2015); inoculum and sequence as determinants | Direction is reliable; exact endpoint is contingent — state as locale-dependent, not deterministic |
| Resource-use efficiency (water, nutrient, metabolic) emerges as a single consequence of the self-reinforcing state, not as separate engineered benefits | Well supported as mechanism; magnitude open | Retention functions (Section 2); self-replenishment vs. passive store (Section 8); Kammann et al. (2015) | Quantifying each efficiency and the maintenance-input floor in the decisive experiment; magnitudes not yet measured |
| Free-energy / self-organization framing of the transition (degradation downhill, building uphill; reinforced stability at a higher, metastable state) | Used illustratively (Figure 2A) | Critical-transition theory as the rigorous basis; dissipative-structure language (Prigogine and Nicolis, 1977) by analogy only | Schematic, not evidence — the energetics are qualitative; the quantitative claim remains the experiment (Section 10) |
References
- Adamatzky, A. (2022). Language of fungi derived from their electrical spiking activity. Royal Society Open Science, 9(4), 211926.
- Akiyama, K., Matsuzaki, K., & Hayashi, H. (2005). Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature, 435, 824–827.
- Amor, D. R., Ratzke, C., & Gore, J. (2020). Transient invaders can induce shifts between alternative stable states of microbial communities. Science Advances, 6(3), eaay8676. https://doi.org/10.1126/sciadv.aay8676
- Babikova, Z., Gilbert, L., Bruce, T. J. A., Birkett, M., Caulfield, J. C., Woodcock, C., Pickett, J. A., & Johnson, D. (2013). Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack. Ecology Letters, 16(7), 835–843.
- Beisner, B. E., Haydon, D. T., & Cuddington, K. (2003). Alternative stable states in ecology. Frontiers in Ecology and the Environment, 1(7), 376–382.
- Borer, B., Tecon, R., & Or, D. (2018). Spatial organization of bacterial populations in response to oxygen and carbon counter-gradients in pore networks. Nature Communications, 9, 769. https://doi.org/10.1038/s41467-018-03187-y
- D'Souza, G., & Kost, C. (2016). Experimental evolution of metabolic dependency in bacteria. PLoS Genetics, 12(11), e1006364. https://doi.org/10.1371/journal.pgen.1006364
- D'Souza, G., Waschina, S., Pande, S., Bohl, K., Kaleta, C., & Kost, C. (2014). Less is more: selective advantages can explain the prevalent loss of biosynthetic genes in bacteria. Evolution, 68(9), 2559–2570. https://doi.org/10.1111/evo.12468
- Dejana, L., Ramírez-Serrano, B., Rivero, J., Gamir, J., López-Ráez, J. A., & Pozo, M. J. (2022). Phosphorus availability drives mycorrhiza-induced resistance in tomato. Frontiers in Plant Science, 13, 1060926. https://doi.org/10.3389/fpls.2022.1060926
- Fujita, H., Yoshida, S., Suzuki, K., & Toju, H. (2025). Alternative stable states of microbiome structure and soil ecosystem functions. Environmental Microbiome, 20, 28. https://doi.org/10.1186/s40793-025-00688-4
- Fukami, T. (2015). Historical contingency in community assembly: integrating niches, species pools, and priority effects. Annual Review of Ecology, Evolution, and Systematics, 46, 1–23.
- Glaser, B. (2007). Prehistorically modified soils of central Amazonia: a model for sustainable agriculture in the twenty-first century. Philosophical Transactions of the Royal Society B, 362, 187–196.
- Glaser, B., Haumaier, L., Guggenberger, G., & Zech, W. (2001). The "Terra Preta" phenomenon: a model for sustainable agriculture in the humid tropics. Naturwissenschaften, 88, 37–41.
- Holling, C. S. (1973). Resilience and stability of ecological systems. Annual Review of Ecology and Systematics, 4, 1–23.
- Jeffery, S., Abalos, D., Prodana, M., Bastos, A. C., van Groenigen, J. W., Hungate, B. A., & Verheijen, F. (2017). Biochar boosts tropical but not temperate crop yields. Environmental Research Letters, 12(5), 053001. https://doi.org/10.1088/1748-9326/aa67bd
- Jeffery, S., Verheijen, F. G. A., van der Velde, M., & Bastos, A. C. (2011). A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agriculture, Ecosystems & Environment, 144(1), 175–187.
- Joseph, S., Kammann, C. I., Shepherd, J. G., Conte, P., Schmidt, H.-P., Hagemann, N., et al. (2018). Microstructural and associated chemical changes during the composting of a high-temperature biochar: mechanisms for nitrate, phosphate, and other nutrient retention and release. Science of the Total Environment, 618, 1210–1223. https://doi.org/10.1016/j.scitotenv.2017.09.200
- Jung, S. C., Martinez-Medina, A., Lopez-Raez, J. A., & Pozo, M. J. (2012). Mycorrhiza-induced resistance and priming of plant defenses. Journal of Chemical Ecology, 38(6), 651–664. https://doi.org/10.1007/s10886-012-0134-6
- Kammann, C. I., Schmidt, H.-P., Messerschmidt, N., Linsel, S., Steffens, D., Müller, C., Koyro, H.-W., Conte, P., & Joseph, S. (2015). Plant growth improvement mediated by nitrate capture in co-composted biochar. Scientific Reports, 5, 11080. https://doi.org/10.1038/srep11080
- Karst, J., Jones, M. D., & Hoeksema, J. D. (2023). Positive citation bias and overinterpreted results lead to misinformation on common mycorrhizal networks in forests. Nature Ecology & Evolution, 7, 501–511. https://doi.org/10.1038/s41559-023-01986-1
- Kiers, E. T., Duhamel, M., Beesetty, Y., Mensah, J. A., Franken, O., Verbruggen, E., Fellbaum, C. R., Kowalchuk, G. A., Hart, M. M., Bago, A., Palmer, T. M., West, S. A., Vandenkoornhuyse, P., Jansa, J., & Bücking, H. (2011). Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science, 333(6044), 880–882. https://doi.org/10.1126/science.1208473
- Kim, J.-S., Sparovek, G., Longo, R. M., De Melo, W. J., & Crowley, D. (2007). Bacterial diversity of terra preta and pristine forest soil from the Western Amazon. Soil Biology and Biochemistry, 39(2), 684–690. https://doi.org/10.1016/j.soilbio.2006.08.010
- Kuzyakov, Y., & Blagodatskaya, E. (2015). Microbial hotspots and hot moments in soil: concept & review. Soil Biology and Biochemistry, 83, 184–199. https://doi.org/10.1016/j.soilbio.2015.01.025
- Kuzyakov, Y., Subbotina, I., Chen, H., Bogomolova, I., & Xu, X. (2009). Black carbon decomposition and incorporation into soil microbial biomass estimated by 14C labeling. Soil Biology and Biochemistry, 41(2), 210–219.
- Kéfi, S., Holmgren, M., & Scheffer, M. (2016). When can positive interactions cause alternative stable states in ecosystems? Functional Ecology, 30(1), 88–97. https://doi.org/10.1111/1365-2435.12601
- Lehmann, J., & Joseph, S. (Eds.). (2015). Biochar for Environmental Management: Science, Technology and Implementation (2nd ed.). Routledge.
- Lehmann, J., & Kleber, M. (2015). The contentious nature of soil organic matter. Nature, 528, 60–68.
- Maillet, F., Poinsot, V., André, O., Puech-Pagès, V., Haouy, A., Gueunier, M., et al. (2011). Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature, 469, 58–63.
- May, R. M. (1977). Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature, 269, 471–477.
- Miller, M. B., & Bassler, B. L. (2001). Quorum sensing in bacteria. Annual Review of Microbiology, 55, 165–199.
- Morris, J. J., Lenski, R. E., & Zinser, E. R. (2012). The Black Queen Hypothesis: evolution of dependencies through adaptive gene loss. mBio, 3(2), e00036-12. https://doi.org/10.1128/mBio.00036-12
- Pineda, A., Kaplan, I., Hannula, S. E., Ghanem, W., & Bezemer, T. M. (2020). Conditioning the soil microbiome through plant–soil feedbacks suppresses an aboveground insect pest. New Phytologist, 226(2), 595–608. https://doi.org/10.1111/nph.16385
- Pozo, M. J., & Azcón-Aguilar, C. (2007). Unraveling mycorrhiza-induced resistance. Current Opinion in Plant Biology, 10(4), 393–398.
- Prigogine, I., & Nicolis, G. (1977). Self-Organization in Nonequilibrium Systems. Wiley.
- Raaijmakers, J. M., & Mazzola, M. (2016). Soil immune responses. Science, 352(6292), 1392–1393. https://doi.org/10.1126/science.aaf3252
- Redfield, R. J. (2002). Is quorum sensing a side effect of diffusion sensing? Trends in Microbiology, 10(8), 365–370.
- Scheffer, M., Carpenter, S., Foley, J. A., Folke, C., & Walker, B. (2001). Catastrophic shifts in ecosystems. Nature, 413, 591–596.
- Schink, B. (1997). Energetics of syntrophic cooperation in methanogenic degradation. Microbiology and Molecular Biology Reviews, 61(2), 262–280.
- Schmidt, H.-P., Kammann, C., Hagemann, N., Leifeld, J., Bucheli, T. D., Sánchez-Monedero, M. A., & Cayuela, M. L. (2021). Biochar in agriculture – A systematic review of 26 global meta-analyses. GCB Bioenergy, 13(11), 1708–1730. https://doi.org/10.1111/gcbb.12889
- Schmidt, M. J., et al. (2023). Intentional creation of carbon-rich dark earth soils in the Amazon. Science Advances, 9(38), eadh8499. https://doi.org/10.1126/sciadv.adh8499
- Schmidt, M. W. I., Torn, M. S., Abiven, S., Dittmar, T., Guggenberger, G., Janssens, I. A., Kleber, M., Kögel-Knabner, I., Lehmann, J., Manning, D. A. C., Nannipieri, P., Rasse, D. P., Weiner, S., & Trumbore, S. E. (2011). Persistence of soil organic matter as an ecosystem property. Nature, 478(7367), 49–56. https://doi.org/10.1038/nature10386
- Slocum, M. P., Barton, R. B., Wagner, S. J., Lehmann, J., Solomon, D., Yeboah, E., & Plante, A. F. (2026). Elevated organic carbon in African Dark Earths is not exclusively attributable to pyrogenic organic matter. Science of the Total Environment, 1016, 181481. https://doi.org/10.1016/j.scitotenv.2026.181481
- Smith, S. E., & Read, D. J. (2008). Mycorrhizal Symbiosis (3rd ed.). Academic Press.
- Solomon, D., Lehmann, J., Fraser, J. A., Leach, M., Amanor, K., Frausin, V., Kristiansen, S. M., Millimouno, D., & Fairhead, J. (2016). Indigenous African soil enrichment as a climate-smart sustainable agriculture alternative. Frontiers in Ecology and the Environment, 14(2), 71–76. https://doi.org/10.1002/fee.1226
- Song, Y. Y., Zeng, R. S., Xu, J. F., Li, J., Shen, X., & Yihdego, W. G. (2010). Interplant communication of tomato plants through underground common mycorrhizal networks. PLoS ONE, 5(10), e13324.
- Takeuchi, N., Fullmer, M. S., Maddock, D. J., & Poole, A. M. (2024). The Constructive Black Queen hypothesis: new functions can evolve under conditions favouring gene loss. The ISME Journal, 18(1), wrae011. https://doi.org/10.1093/ismejo/wrae011
- Todman, L. C., Fraser, F. C., Corstanje, R., Harris, J. A., Pawlett, M., Ritz, K., & Whitmore, A. P. (2018). Evidence for functional state transitions in intensively-managed soil ecosystems. Scientific Reports, 8, 11522. https://doi.org/10.1038/s41598-018-29925-2
- Tsoi, R., Wu, F., Zhang, C., Bewick, S., Karig, D., & You, L. (2018). Metabolic division of labor in microbial systems. Proceedings of the National Academy of Sciences, 115(10), 2526–2531.
- van der Heijden, M. G. A., Klironomos, J. N., Ursic, M., Moutoglis, P., Streitwolf-Engel, R., Boller, T., Wiemken, A., & Sanders, I. R. (1998). Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature, 396, 69–72. https://doi.org/10.1038/23932
- Wang, G., van der Putten, W. H., Klironomos, J., Zhang, F., & Zhang, J. (2025). Steering plant-soil feedback for sustainable agriculture. Science, 389(6758), eads2506. https://doi.org/10.1126/science.ads2506
- Wang, J., Xiong, Z., & Kuzyakov, Y. (2016). Biochar stability in soil: meta-analysis of decomposition and priming effects. GCB Bioenergy, 8(3), 512–523.
- Waters, C. M., & Bassler, B. L. (2005). Quorum sensing: cell-to-cell communication in bacteria. Annual Review of Cell and Developmental Biology, 21, 319–346.
- Weller, D. M., Raaijmakers, J. M., Gardener, B. B. M., & Thomashow, L. S. (2002). Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annual Review of Phytopathology, 40, 309–348.
About this companion
This is the plain-language companion to the technical paper of the same name. It follows that paper section by section and, as far as it can, paragraph by paragraph, in the same order, so the two can be read side by side. It can be read on its own as well. One thing to hold onto from the start: this paper proposes an explanation, it does not claim to have proven one. It gathers a great deal of established science to show why the explanation is worth taking seriously, and it is careful, throughout, to mark the line between what is known and the one big experiment that has not yet been run. This plain version keeps that line exactly where the technical paper draws it. The scientific sources are all in the technical paper.
Abstract (the whole argument in short)
Terra preta, the dark earth of the Amazon, is more fertile, more alive, and longer-lasting than the worn-out tropical soil around it, and it has stayed that way for centuries after the people who made it stopped tending it. This paper offers a single explanation for that, and assembles the science that motivates it: that a tough, sponge-like skeleton of charcoal acts as a shared, multi-purpose home for soil life, and that adding such a skeleton, along with a complete set of minerals and a starter culture of microbes, can let an ordinary soil mature into a genuinely different kind of system. The argument has four parts.
First, the charcoal skeleton supplies living space, the ability to grip nutrients, reservoirs of water and food, a place for chemical signals to linger, and shelter, all through different sizes of pore and different surface chemistry that come online on different timescales. Second, the resulting jump in how densely and how closely the microbes are packed switches on a kind of community economy, in which being close together is the thing that turns a shared division of labor from a losing proposition into a winning one; and over a much longer, evolutionary timescale, the microbes shed genes for things their neighbors already provide, which deepens their dependence on one another and makes the whole arrangement more stable rather than less. Third, the self-reinforcing loops created by that economy are exactly the precondition, in ecological theory, for a system to have a second stable state; soil communities are increasingly inferred to have such alternate states with a "stickiness" that makes the idea of an ecological tipping point rigorous rather than poetic. Fourth, fungi act as the system's connecting network, moving its minerals, trading back and forth with their plant partners, and priming plant defenses, with all of these jobs getting stronger as the fungal network gets denser. On this telling, the soil’s staying power is really two kinds of lasting working at once: a tough carbon scaffold that endures on its own, doing nothing, and a living community that holds fertility up only while it is fed, the scaffold a durable home for the engine and the engine supplying what the dead scaffold cannot. The same arrangement also predicts a distinctive thrift, tight recycling of nutrients, a water-saving economy, and fertility that largely renews itself under continued harvest, all of it falling out of the mechanism rather than bolted on.
Every part the explanation needs is independently well established across five different fields of science; their agreement on one blueprint is what makes the idea worth testing, though it does not by itself prove that the parts actually assemble into the whole in a real soil. The explanation is coherent and testable. What remains unproven, and what would turn it from a strong idea into a discovery, is a single experiment: a measured, deliberate crossing of a soil into the high-fertility state, with the tell-tale "stickiness" that marks a genuine new equilibrium.
1. Introduction: a soil that runs itself
The real puzzle of terra preta is not that it is fertile. Plenty of soils are fertile while you keep feeding them. The puzzle is that terra preta stayed fertile, alive, and structurally distinct for centuries to thousands of years after people stopped adding anything to it, sitting in the middle of a landscape of heavily leached, nutrient-poor soils that do not behave that way. A soil that holds a high-functioning state with no upkeep is behaving less like a fertilized field and more like a system that has settled into a different equilibrium.
The idea offered here is that this difference is produced, and then held in place, by a physical object: a tough, sponge-like skeleton of charcoal. Supplied alongside a complete set of minerals and a starter culture of microbes, that skeleton does one specific and consequential thing: it adds an enormous amount of structured living space and reactive surface. That addition raises how densely the microbes pack and, more importantly, how close together they are, to the point where the community is no longer held back by lack of room, by shared resources drifting away, or by losing its partners. Past a threshold, with resources plentiful enough that nothing is the bottleneck, the system does not just hold more of the same life. It crosses into a different kind of organization that can both assemble itself and stay assembled, and it is that self-sustaining, higher-functioning state that this paper identifies with terra preta.
The central claim can be put in one line: the conditioned charcoal skeleton is the keystone of the whole terra preta package. One material, charcoal of enough quantity, aged to grip nutrients well, loaded with a complete mineral profile, and seeded with life, is the controlling ingredient from which the soil's other properties follow, rather than a checklist of separate traits each installed by hand. Those properties come in two kinds of payoff, on two clocks. The first is immediate and physical: better crumb structure and tilth, water held against drought, and resistance to nutrients washing away, all of which a high-surface, high-grip material delivers the moment it enters the ground, with no biology and no waiting. The second is slower and biological: the self-renewing fertility that lasts under cropping, the holding-down of disease, and the organized microbial complexity that emerge as ordinary soil physics and biology work on that skeleton over time. The strong form of the claim is that this second payoff genuinely emerges, the result of feedbacks running on the right material rather than a set of separately engineered functions, and that it is therefore reproducible in principle by conditioning charcoal to the same recipe, which reframes the common belief that terra preta cannot be deliberately remade as a solvable problem of conditioning rather than an unsolvable mystery. The claim carries one honest limit: that second, biological payoff is reliably there given a good starter culture and enough time, but it is not instant and it is not indifferent to how things begin, because the order in which a community assembles means the same material can be colonized along more than one path.
(The paper also states, up front, that the synthesis rests on established science and that the integrated result, a deliberate, measured crossing into the self-sustaining state, remains the open experiment; it is explicit about where that boundary lies.)
2. The charcoal skeleton as a multi-purpose shared home
The starting point is a number. Charcoal can present an enormous amount of surface area for its weight, anywhere from about twenty to several hundred square meters per gram, and more for activated carbons. The soils where charcoal matters most are the opposite: sand has almost none, silt very little, and the non-swelling clay that dominates the weathered tropical soils of the Amazon comparison only ten to thirty square meters per gram. So adding even a modest fraction of charcoal to a low-surface soil can raise its total surface area a great deal, by several times in many soils and by close to tenfold in sandy, weathered, low-clay ones, which is exactly the kind of soil terra preta arose in.
An important refinement: "surface area," as the lab measures it with gas, is not the same as space a microbe can actually live in. Most of that measured area is in pores far too small for a cell. The space microbes truly occupy is the bigger, micron-scale pores, sized to fit cells and fungal threads. So a raw surface-area number overstates the actual living quarters, often several times over.
That distinction does not weaken the idea; it sharpens it into a two-job picture in which the same material does different things through different pore sizes. The bigger pores are housing: where cells and fungal threads live, and where they are sheltered from the protozoa that graze on them, so the density gain is real and not just theoretical. The finer surface is pantry and switchboard: it soaks up and holds water, dissolved food, gases, mineral nutrients, and the chemical signals the community uses, keeping them close instead of letting them wash away. That second job is the physical basis for the community economy of Section 3, and the fact that one material does both jobs is the heart of the "multi-purpose home" idea.
A frequently muddled point concerns the soil's ability to grip nutrients, its cation exchange capacity, the static-cling-like hold that keeps positively charged nutrients from leaching out. That grip does not come from bare surface area. It comes from oxygen-bearing chemical groups on the carbon surface. And here is the catch: making charcoal hot builds surface area but burns those gripping groups off, so surface area and fresh grip tend to pull in opposite directions, with grip strongest in lower-temperature char. At first this looks like a problem for any "more surface, more function" story, but two things resolve it and, in doing so, strengthen the case that terra preta is grown.
First, living space, surface area, and nutrient grip are set by three different things, so they are partly separate dials, not one slider. The big living-space pores come mostly from the original cell structure of the plant that was charred; the fine surface area comes from high-temperature charring; the nutrient grip comes from low-temperature and, crucially, aged surface chemistry. And since how much you apply is a free extra dial, there is no forced trade of grip against surface area for the system as a whole: a heavier dose of a lower-temperature, higher-grip char can supply both living space and grip, at the cost of more material rather than lost function.
Second, and more interesting, is timing. The physical side, the pores and surface, is fixed the moment the charcoal is made. But much of the surface chemistry, and therefore much of the nutrient grip and a large part of the biological function, develops over time as the char weathers and its surface oxidizes, the way a cast-iron pan seasons with use. The housing is ready on day one; the gripping economy and the biological function ripen over months to years. Different functions come online on different clocks. This staggered ripening is not a complication to explain away. It is the very reason terra preta is something grown rather than mixed: if every function switched on at once when you applied the char, there would be nothing to wait for, and the well-known observation that dark earth develops over time would make no sense.
One consequence of all that grip and sponge is that it stops nutrients from washing away, and that matters most exactly where terra preta arose. In a high-rainfall tropical landscape over low-grip soils, soluble nutrients from rain and irrigation are flushed below the roots before plants can use them; the charged, sponge-like skeleton catches and holds those nutrients against that flow, so fertility added to the soil stays in the soil. A simple drip-through column is one of the most direct ways to test this (Section 10).
The same holding power extends to water, on two levels. Physically, the pore structure soaks up and holds water, raising the soil's water-holding capacity, most noticeably in coarse, low-clay soils. Biologically, the dense community the skeleton supports secretes slimes and gels and builds crumb structure that together hold moisture and slow drying, so a larger, more active population of microbes becomes, in effect, a second water reservoir built on top of the first. The drought buffer is doubled, once by the physical cavities and again by the life those cavities house. (To be precise, the biological water is held in the secreted gel and the improved structure, not stored inside the microbes themselves, whose volume is tiny.)
These holding properties carry a corollary that is easy to miss and that matters for how the skeleton should be prepared and tested. Fresh, uncharged char is none of these things yet: its gripping sites and sponge surfaces are empty, so when it first enters a soil it pulls nutrients and water out of solution and holds them, behaving for a while as a net taker that demands from its surroundings rather than a giver that contributes. Only once those sites are loaded, the surface aged, and the life established does the same material become a net contributor, releasing and cycling what it holds. This switch from taker to giver is the practical face of the staggered ripening above, and it is why the manner and the timing of preparation, not the bare presence of char, decide whether the skeleton helps or harms in its first seasons. It is also the most likely reason short trials of fresh, unseeded char give such inconsistent results, a point taken up in Section 8.
3. The crowd economy: density, division of labor, and dependence
If the charcoal skeleton raises density and closeness, the question becomes what changes when a microbial community gets dense and tightly packed. The microbiology literature answers with several well-supported, named mechanisms that together describe a self-reinforcing economy.
Switching on at a crowd size. Quorum sensing is the classic case of a microbial community changing its behavior once it gets dense enough. Cells release signaling molecules whose concentration rises as the population grows; above a threshold, the population switches on group behaviors like forming biofilms and coordinating metabolism. So the claim that "past a certain density the system flips into a new mode" is not speculation; it is a central idea in bacterial social life. One honest note: how sharp that threshold is gets debated, and single-cell studies suggest the switch can be gradual rather than a hard cutoff, so the careful phrasing is "a threshold-like shift into a new mode," which is all the argument needs.
Tight quarters lower the crowd size needed. Beyond plain density, there is good support for the idea that these same systems respond to physical confinement, so that even a lone cell in a small enough enclosed space acts as if it were in a crowd, because its signals cannot drift away. Inside a porous charcoal skeleton, a cell in a small pore behaves as though surrounded by a dense crowd. The structured home does not just hold more cells; it lowers the crowd size needed to trigger group behavior. That is a direct mechanical link between pore space and the community economy.
Division of labor, and the closeness that ties the whole argument together. Splitting a long chemical task across specialists lightens the load on each cell, and models predict this can raise the community's output and let more species share the same resources. The crucial and often-omitted result is that division of labor is not automatically more efficient. Splitting a job across cells adds a transport cost, because the half-finished products have to travel between them, and a careful analysis shows a two-strain team actually makes less than a single all-in-one organism of the same ability, unless specializing improves efficiency enough to beat that transport cost. And what sets the transport cost is distance. In a watery, well-mixed setting, the shared products leak away between partners and division of labor can lose; pack the partners a few microns apart in structured space and the transport cost collapses, tipping the balance so the economy wins. This is the point where the paper's two main claims meet: the closeness supplied by the charcoal skeleton is the very thing that flips division of labor from a loss into a gain. The "real estate" claim and the "efficiency" claim are not two assertions but one mechanism.
A second, slower clock: shedding what your neighbors provide. The mechanisms above act on everyday, physiological time and need no genetic change. A further process acts on a different clock, an evolutionary one that changes which genes the microbes carry, and rather than competing with the closeness economy it compounds it. The idea (called the Black Queen Hypothesis, after the card game where you avoid the costly queen) is that an organism gains by dropping a costly function that leaks a public good into the neighborhood, letting its neighbors supply it, the way a household might give up its own well once the town has a water main. This drives genome streamlining and produces deep mutual dependence. Because it needs mutation and selection across generations, it is not what establishes the high-functioning state within a single season; it enters from two directions. First, the microbes that seed a soil are already dependent and cross-reliant, since gene-shedding is common in nature, which is part of why most soil microbes are so hard to grow alone, so the closeness economy draws on a deep, ready-made web of dependence from day one. Second, more gene-shedding continues within the established community, fast in generations but slow in real time because most soil cells grow slowly, so it plays out over the soil's long life, and as it proceeds the dependence tightens and the community's "valley" deepens. A modeled extension suggests this can be creative as well as subtractive, with freed-up capacity allowing new functions to evolve, so the economy compounds rather than merely levels off. The honest boundary is that this second clock, especially its creative form, is an evolutionary argument and partly a modeled one; it is offered as a reinforcement that entrenches the state over the soil's life, not as the immediate engine of the crossing, which remains the closeness economy above.
Multiplying, not adding. These benefits are not separate perks to be summed. Density enables specializing; specializing lowers each cell's cost; lower cost supports higher density and tighter coupling. The terms feed one another, which is what produces a threshold rather than a smooth ramp. This is the flywheel, turning on everyday time, with the gene-shedding clock turning slowly beneath it, so a state set up by the fast loop is deepened by the slow one. Section 4 shows that this kind of self-reinforcing loop is exactly the formal requirement for the regime shift the paper proposes.
The boundary of this section is that all of the above is established as mechanism, supported by theory and by lab and synthetic-community experiments. What it does not by itself show is that a real soil crosses into a distinct, higher-order state. It shows that every gear the flywheel needs is real.
4. Crossing the threshold: tipping points and the two-valley picture
The intuitive idea of an ecological "escape velocity," a hump the community must clear before a higher organization becomes self-sustaining, has a rigorous, long-standing form in ecology: the tipping point into an alternative stable state.
The theory holds that an ecosystem can rest in two or more stable states under identical outside conditions; when a disturbance pushes it past a critical point, it responds sharply and disproportionately and shifts into the other state. The "stays assembled" half of the intuition corresponds to "stickiness" (hysteresis): once you have crossed the tipping point, putting conditions back the way they were does not put the system back, so a small change can produce a lasting one. The familiar picture is a ball in a valley. An "escape velocity" is the push needed to roll the ball over a ridge into the next valley, not a runaway to infinity; once in the new valley, knocks roll back toward it, which is the rigorous form of the self-maintenance the idea emphasizes.
Two things make this more than an analogy here. First, a self-reinforcing loop is necessary but not sufficient for a second stable state, because whether a given loop actually opens a second valley depends on its form and strength, not merely on its being present. So the closeness economy of Section 3 and the multi-purpose surface of Section 2 are not separate conveniences; they are the self-reinforcing loops without which a second valley cannot exist.
Second, a growing body of evidence puts these transitions in microbial communities specifically, and increasingly in soil, not just in lakes and forests. An analysis of more than fifteen hundred farm-soil samples found that microbial community structure sorts into several alternative stable states separated by apparent tipping points, with the states differing in how much crop disease they carry. In controlled microbial communities the switch has been driven and watched directly: a brief disturbance, including a temporary change in acidity, can flip the community into a lasting alternative state that holds after the disturbance is gone. Soil function shows the same signature under drought stress, where a mild disturbance lets the community return to normal but an intense one pushes it into an alternative state that persists. That persistence after the cause is removed is the field-scale form of the self-sustaining state this paper proposes. The soil evidence so far shows that such states exist and that switches happen; tracing a full "stickiness" loop in soil, rather than inferring the tipping points, is the harder measurement still to be done.
The most directly relevant home for the argument is restoration ecology. For degraded systems with alternative stable states, the road back is not the mirror image of the road down, and the feedbacks make restoration hard; degraded systems get trapped in their state, so simply reversing the original cause is not enough to bring function back. In the field's own terms, this is exactly the present argument: a worn-out soil is not merely low on inputs but is held in a low-fertility valley by its own feedbacks, and cannot be coaxed out by adding a little at a time. The rigorous justification for a comprehensive, all-at-once intervention (carbon skeleton, complete minerals, and starter culture delivered together as one over-the-threshold push) is exactly this: half-measures relax back, whereas one threshold-crossing push can reorganize the system into the higher valley, where it then stays. The clearest way to say what that all-at-once push must be is that a living soil community of this kind behaves like an engine, not a pile of parts. The carbon home, the full set of minerals, and the living starter all have to come together and reach a minimum of working order at the same time before the self-feeding loop will run on its own, the same way an engine with no fuel, or no spark, or no compression does not run a little, it does not run at all. Adding a bit at a time is like turning the engine over with no fuel in the line; the complete, all-at-once charge is what lets it catch and keep running. The same picture explains the messy biochar record in Section 8.
The idea that ties the paper together is that the charcoal skeleton is the durable, non-living anchor of the stabilizing loop. Alternative stable states are usually driven by feedbacks between the living and non-living parts of a system. The skeleton lasts (Section 7), and while it lasts it holds the water, nutrients, signals, and habitat (Section 2) that sustain the dense, interdependent, efficient community (Section 3), which in turn keeps building the crumb structure and mineral-organic bonds that hold the system in the high-fertility valley. The roles are distinct and should not be blurred: the durable carbon is the anchor whose lasting presence keeps the stabilizing loop supplied, so the valley does not erode over time, but the valley itself, and its stickiness, are produced by the self-reinforcing biology the material supports, not by the dead carbon. The carbon makes the state durable; the living feedback makes it a place the system settles into. And here is the careful part: terra preta lasting for centuries after abandonment fits this picture, but it is not by itself proof of a living, self-sustaining state, because durable carbon and passive nutrient-grip would last for centuries with no self-reinforcing biology at all. Endurance has more than one possible cause. What would tell a genuine self-sustaining biological state apart from a durable-but-passive nutrient bank is not that the soil lasts but how it lasts: a passive bank is drawn down as harvests carry nutrients off, whereas a self-renewing community would keep fertility up under continued cropping. That behavioral tell, lasting under harvest rather than lasting at rest, is the deciding test taken up in Sections 8 and 10.
Three boundaries, stated honestly. A sharp response by itself does not prove there are two states, because sharp responses can also follow sharp changes in outside conditions; proving an alternative stable state requires showing the stickiness, two states under the same conditions. The documented soil transitions are mostly downward, into degraded states that are hard to reverse; the deliberate upward crossing into a high-fertility self-sustaining valley is strongly plausible by the symmetry of the theory, and is supported by the dark earths existing as a built high-fertility endpoint, but it has not been shown as a controlled experiment. And a real feedback is necessary but not sufficient; the loops must be strong enough to fold the system's response curve and open a second valley, and showing they are that strong is the work that remains.
5. The fungal layer: the soil's connective tissue, not its mind
Among the parts of the proposed system, the fungal network reaches the furthest, and the right approach is to state its role at full strength and then to say, just as clearly, where the evidence for that role stops. The functions sort into five of differing strength: the network ties the system together, moves its minerals, runs a give-and-take market with its plant partners, primes plant defenses, and carries signals. The first four are well supported. The fifth is partly supported and carries one famous overstatement that has to be set aside.
Tie the system together (well supported). The fungal network is the closest thing soil has to a connecting organ. Fungi run large parts of the soil economy at once: they regulate nutrient cycling, shape which microbes thrive, hold down soil-borne diseases, and glue mineral and organic particles into the crumbs that give soil its structure, while their dead bodies and the carbon they get from plants feed the stable organic-matter pool. This connecting role is measurable for the whole system, not just inferred: in experiments, changing the variety of root-partnering fungi below ground changed the variety, the stability, and the productivity of the whole plant community above. That is the strongest sense in which the network organizes the system, a change in the fungal layer reorganizes the output of the whole. It is best pictured as connective tissue running through every layer of the soil, not a controller sitting on top. (One careful note on carbon: the role is two-sided, since dead fungal matter stores carbon while active fungi can also speed its turnover, so the defensible statement is that fungi govern the carbon balance, not that they only store carbon.)
Move and shuttle minerals (well supported). The network is also the plant's main organ for getting the hardest-to-reach minerals. Fungal threads extend the root's reach enormously and bring back phosphorus, nitrogen, water, and trace elements the root alone cannot reach, including elements pried off mineral surfaces by fungal activity. On a durable, high-grip carbon home, this network is how the system draws fresh mineral stock into biological circulation, the same reach the companion framework develops as a downward mining of the rock below.
Run a give-and-take market (well supported). The exchange between plant and fungus is not a passive leak but a regulated, two-way trade. Plants can detect, sort, and reward the fungal partners that deliver the most phosphorus by sending them more carbon, and the fungi in turn send more nutrients to the roots that send them the most carbon, so cooperation is rewarded and cheating is penalized on both sides. This is a real marketplace with partner choice and consequences, and it is the rigorous content of the intuition that the system "negotiates": sorting and reward, not deliberation. The careful note is that the reward mechanism is real but depends on context, shifting with how hungry the host is and what is on offer rather than running by a fixed rule.
Prime the immune system (well supported, and a literal immune function). There is a named, repeatedly confirmed effect in which colonization by root-partnering fungi gives the plant's immune system a mild wake-up, so the plant is "primed" to respond faster and harder when an attacker arrives, like a vaccine readying a defense. It has been shown across many pests and diseases in many crops. The honest note is that it depends on context; under high-nutrient or high-light conditions the priming can shrink or reverse, so the defensible phrasing is that fungi prime plant immunity under most conditions.
Signal and communicate (mixed, with one overstatement to set aside). The communication claims have to be sorted by strength. The plant-fungus chemical conversation is settled science: under phosphorus stress the plant releases a signal that makes the fungus germinate and branch, and the fungus answers with a signal the plant's receptors recognize, switching on the partnership. Beyond that point the evidence weakens and must be labeled honestly. Individual experiments report that plants linked by a shared fungal network can pass along defense cues, with linked neighbors showing more resistance or releasing defensive scents when a donor is attacked; these results are suggestive, but they are among the single-study findings that a careful review found to be over-cited and not yet robustly established, so they are presented here as contested, not settled. Electrical activity resembling nerve impulses has been recorded traveling in fungal threads; this is a real phenomenon, but reading it as a "language" is unsupported speculation and is not evidence of thinking, and it is treated here as an open frontier at most. The romantic version of all this, that forests are a "wood-wide web" through which old trees consciously share food and warnings with their kin, was formally challenged by a review that found the claims of widespread networks, of reliable resource transfer that helps the receiver, and of parents favoring their offspring to be insufficiently supported or, in the last case, without peer-reviewed evidence at all. That story is set aside, while the chemical conversation, the immune priming, and a cautious, contested plant-to-plant signaling are kept.
The point that ties this layer to the rest of the paper is that these fungal jobs get stronger as the fungal network gets denser, with transfer through the network reported to rise with network density. So if the charcoal skeleton supports a denser fungal network (Sections 2 and 3), then all of these jobs, connecting, gathering, trading, priming, signaling, intensify with that density. The fungal layer is not a separate claim; it is one more set of jobs riding the same surface-area-and-density flywheel.
Plainly, then: the evidence supports a fungal network that ties the system together, gathers and moves its minerals, trades back and forth with its plant partners, and primes their defenses, with all of these strengthening as the network densifies. That is a commanding organizing role, and the paper rests on it. What the evidence does not support, and what this paper does not claim, is that the network is intelligent, that it deliberates, or that it consciously runs the system from above. The organization it provides is the emergent product of distributed function and two-way exchange, the connecting tissue of the soil, not its mind, and stating the role at exactly that strength is what keeps it defensible.
6. Five fields, one blueprint
The strongest support this synthesis can offer is not any single study but a pattern of agreement. Five fields of science that do not usually cite one another each describe a piece of the same blueprint: dense, structured, well-fed communities crossing feedback-driven thresholds into self-sustaining, high-functioning states. When independent fields, studying different systems, each recover a piece of one blueprint, that is good evidence the pieces are real and the blueprint is built from established parts. It is not, by itself, evidence that those parts assemble into the whole in a soil, which is the step Section 10 isolates as the open experiment.
Disease-suppressing soils (microbiology). A suppressive soil is one where a soil-borne disease fails to take hold even though the germ, a vulnerable crop, and good conditions for disease are all present, because of the microbial community; it has been studied for decades and framed as a kind of soil immune response. It comes in two forms that map onto this paper's claims: general suppression, driven by sheer microbial mass, an inherent property of the soil that cannot be transferred (the density claim, as community-wide immunity), and specific suppression, due to particular microbes and transferable by mixing a little suppressive soil into a vulnerable one, and destroyed by heat (a near-demonstration of transferring the state by seeding, which is the intervention model). The careful note is that general suppression is a correlation rather than a switchable state, and that suppressive soils are often hard to predict.
Living together in tight spaces (soil physics). Experiments in tiny artificial pore networks show two bacterial species, one needing oxygen and one not, settling into different regions along opposing food and oxygen gradients and coexisting in structured space in a way that is impossible in a well-mixed broth. This is the "skyscraper versus open field" intuition shown directly: structured space supports coexistence that mixing destroys. Soil is also understood to organize into hot-spots of locally intense activity, at places like root surfaces, decaying litter, and pore walls, with the size of those hot-spots set by how far food and signals diffuse through the pores, which fuses the surface argument of Section 2 with the economy of Section 3.
Who-gets-there-first (community ecology). The order and timing of arrival cause lasting differences in which community forms, and can produce alternative stable states. Which valley a community lands in can therefore depend on who arrives first, which sharpens the intervention model: it may not be enough to add resources and life; the desired community may need to be established first and fast, so it claims the space before the degraded community reasserts itself. The honest crack is the live debate over how much of assembly is predictable versus chancy, and the fact that some disturbed systems drift back toward their original state, so a new state has to be genuinely self-sustaining, not just briefly different.
Plants and soil shaping each other (agroecology). Plants condition the soil and the soil conditions the next plants, a feedback now being developed deliberately as a tool. A proof-of-concept showed that steering the soil community this way can give a crop above-ground resistance to an insect pest, and a recent synthesis argues for using these principles to restore soil function in farming. This connects suppressive soils, who-gets-there-first effects, and the present idea: conditioning produces a protective community that can be steered by seeding. The crack is that the feedback can be negative as well as positive, and that how to steer communities toward their helpful functions is still largely unknown.
Living dark earths today (human-made soils). Terra preta is not unique to the Amazon, and, crucially, the method is not entirely lost. West African communities make carbon-rich, high-fertility dark earths today, much like Amazonian terra preta, by piling ash, char, bones, and organic waste into kitchen gardens and middens. These soils hold much more organic and charcoal carbon, more available phosphorus, and a stronger nutrient grip than the soil next to them. The honest crack is the lingering debate over intention: it may well be that Amazonian terra preta was not deliberately engineered for fertility on a large scale, so the intentional, engineered-at-scale version is the thing still to be shown, which is again the open experiment.
The unifying observation is the agreement itself. Microbiology, soil physics, community ecology, ecosystem theory, and restoration ecology each, in their own systems, describe a piece of one density-and-feedback-driven crossing into a self-sustaining high-functioning state. That agreement is what makes the idea credible and worth testing, independent of any single source; what it does not do is stand in for the whole demonstration, which remains the open experiment of Section 10.
7. The carbon question: how long it lasts, and the limits of "forever"
The mechanism in Sections 4 and 6 depends on the charcoal skeleton lasting long enough to anchor the stabilizing loop. That raises two questions: how durable the skeleton really is, and whether anything in soil is truly permanent.
The modern understanding of soil carbon directly supports the framing used here. The field has largely dropped the idea that certain molecules are inherently un-rottable; lasting is now understood as a property of the situation, physical protection, being locked to mineral surfaces, the local absence of the right organisms or enzymes, oxygen, moisture, rather than a fixed trait of the molecule. Charcoal carbon fits this picture as an unusually durable but still finite material. Isotope and incubation studies split it into a small quick-rotting fraction that disappears within months and a dominant durable fraction that lasts centuries. The dense, ring-structured core is that durable backbone.
There are three different fates for the skeleton, and only one looks like simple rearrangement. The quick fraction is genuinely burned off to carbon dioxide and lost (more of it in lower-temperature char). The surface is also oxidized, but productively, building the oxygen-bearing groups that grow the nutrient grip over time. The dense core mostly lasts, slowly, for centuries. For building a fertile soil, the early loss is modest and much of the breakdown is itself the ripening: fragmentation exposes fresh surface, surface oxidation builds grip, the quick carbon feeds the early microbial bloom, and core fragments bond with minerals into durable mineral-organic complexes. For carbon-storage accounting, by contrast, a lower-temperature char does cost something, because permanence rewards the most durable material. One local note: charcoal may turn over faster in tropical soils than global averages suggest, which would mean faster ripening but shorter permanence there.
A further part of the skeleton's contribution to durable carbon is indirect. A large share of the organic carbon stored in the dark earths is not charcoal at all, and recent work finds their carbon enrichment cannot be put down to charcoal alone. Consistent with the "it's the situation, not the molecule" view, the durable carbon and the mineral surfaces it helps organize plausibly protect ordinary decayed organic matter sitting alongside it, extending its lifetime beyond what it would have on its own. So the durable-carbon benefit is partly the skeleton's own toughness and partly the protection it lends the non-charcoal material around it, which also means a complete soil-building input includes the rottable organic matter that pool is built from, not the skeleton alone.
On the broader claim that nothing is permanent given enough time and fuel, the honest position is that this is more right than wrong, but the absolute version overshoots, and where it breaks is instructive. Time wears down "slow" barriers, so slow reactions eventually proceed; time does nothing to "uphill" ones. A transformation that is energetically uphill at ordinary conditions will not proceed by microbes no matter how much time passes or how much unrelated energy is lying around nearby, unless it is hitched to a sufficiently downhill partner reaction, and there is a minimum energy payoff a cell can pocket, below which no organism bothers and so no enzyme ever evolves, like a minimum wage below which no one takes the job. This is why "given enough fuel" misframes it: the organism has to earn energy from the target reaction itself, not from random calories nearby. Fully oxidized minerals offer nothing to burn for energy, and "forever chemicals" (PFAS) are a clean modern example of a material that resists being broken down for want of an energy handle. The densest carbon, including the core of the skeleton, offers few accessible edges to attack, which is exactly why that fraction lasts longest.
The useful conclusion is the one the field itself adopts: "permanent versus breakable" is the wrong question, and "how fast does it break down under these specific conditions" is the right one. Under that framing the philosophical point holds (almost nothing is truly permanent) while the absolute claim does not (a few things are effectively inert for lack of energy or a handle, and an uphill reaction is a wall time does not sand down). The practical payoff is that soil-carbon lifetimes span an enormous range, and that spread is the whole value: a skeleton that lasts centuries while slowly feeding the system is what makes the mechanism work, and it is exactly the caveat that must attach to any "permanent carbon storage" claim. The craft is not making the skeleton permanent or breaking it, but tuning its rate.
One implication of this durability belongs to the argument that follows and is worth stating plainly. Precisely because the skeleton and its passive grip last on their own for centuries, the mere endurance of a dark earth cannot be evidence that a living, self-reinforcing community is holding it in place, since the carbon explains the long life with no help from biology. The biological claim of this paper therefore does not rest on persistence as such. It rests on how a soil persists, the distinction developed in Sections 8 and 10.
8. The competing explanation, and the messy biochar record
An idea like this earns its place only by facing the most economical rival explanation and the mixed real-world record honestly, not by collecting confirmations. For terra preta and for biochar there is a strong rival and a genuinely uneven track record, and both are taken up here directly, because both, read carefully, narrow the claim rather than defeat it.
The rival explanation is non-biological, and it is the simplest default. On this account the fertility and the durability of dark earth are products of chemistry, not of any change in biological organization: big stocks of phosphorus and calcium from bone, ash, and waste; a strong nutrient grip that holds cations; a raised, buffered acidity; and a durable carbon skeleton that lasts for centuries. The West African dark-earth data fit this reading, since those soils carry several to many times more available phosphorus, a stronger grip, and a friendlier pH than their neighbors, and the biochar review record pins a large part of biochar's yield benefit on a simple liming (acid-neutralizing) effect rather than anything subtler. This reading is reinforced by how these soils are now understood to have formed in the first place: people built them over many generations by piling up nutrient-rich leftovers, fish and animal bone, ash, charcoal, and household refuse, in and around where they lived, whether on purpose to improve the ground or simply as the residue of living there. That formation history is the strongest version of the chemistry account, a nutrient subsidy paid in slowly across centuries rather than a single act of engineering, and this paper’s claim that the state can be set up quickly and then largely keep itself going has to be measured against that long build-up, not assumed past it. The honest position is that this account is real and may explain a large share, perhaps the majority, of what makes dark earth fertile and durable. A responsible version of this paper's idea does not deny the chemistry; it states precisely what it adds to it.
What it adds is an answer to two things the chemistry alone does not explain. The first is self-renewal under harvest. A nutrient bank with a strong grip is still a fixed store, and a store is drawn down as harvests carry nutrients off, so passive chemistry predicts slow decline, not the keeping-up of fertility under continued use. A living community that fixes nitrogen, frees phosphorus, and rebuilds organic matter is the kind of system that could keep a soil renewed rather than merely well-stocked, and self-renewal, not storage, is what would set a soil in a self-sustaining state apart from a rich but finite bank. The second is the shape of the response. Add-and-it-scales chemistry predicts that benefit rises smoothly with input and reverses when you stop, whereas the regime-shift claim predicts a threshold, a disproportionate response once it is crossed, and persistence afterward. Terra preta is also found to host distinct, lasting microbial communities, different from the soil next door, which fits a distinct biological state, though on its own it cannot show the biology is a cause rather than a result of the chemistry. The two are separated not by assertion but by measurement: whether the high-functioning state lasts under continued harvest, whether it shows threshold-and-stickiness behavior rather than smooth reversibility, and whether phosphorus ends up in biologically re-releasable forms rather than passively stuck or lost. Those are the experiments of Section 10, designed to test the biological claim against the chemistry rather than to assume it.
The biochar track record must be read with the same honesty, because at first glance it looks unfavorable to any strong claim. Across reviews the average yield benefit is modest, around ten to thirteen percent, and highly variable, with many no-effect and some negative results. Part of that variability is publication-sensitive, and at least one analysis found a headline benefit flip sign after correcting for the tendency to publish positive results. None of that is brushed aside here. But the same record carries a pattern that is the opposite of discouraging for this idea: the benefit is concentrated in acidic, sandy, low-grip, weathered soils in tropical climates, with large gains where the starting acidity is high and little or no effect in neutral, fertile, temperate soils. That is not random scatter. It is the dark-earth setting itself, the soils where the proposed mechanism should matter most and where ordinary ground has the least to begin with.
The rest of the scatter is, to a large degree, an artifact of how biochar has been tested, and this is where the preparation argument of Section 2 becomes decisive rather than rhetorical. Most trials apply fresh, uncharged, unseeded char and measure the result over one or two seasons. By the taker-to-giver argument, that is exactly the material, and exactly the window, in which char should perform worst, because an uncharged skeleton first pulls nutrients and water out of solution before it gives them back. The prediction is borne out directly: in a controlled comparison, untreated biochar cut plant growth to sixty percent of the no-char control, while the same biochar, composted-and-charged with nutrients before use, raised growth by up to three hundred percent in the same poor soil, traced to captured nitrogen and phosphorus held against leaching and released slowly. Fresh char and charged char are, in effect, two different materials, and a literature that has mostly tested the first cannot be read as a verdict on the second.
This is the engine picture from Section 4, applied to the record. Most biochar trials were not weak tests of the whole engine; they were tests of an engine with parts missing, a bare skeleton, or a skeleton plus a splash of acid-correction, run for a season or two. That is the engine turned over with no fuel and no spark, and its failure to start says nothing about the assembled engine. Running an engine with missing parts and no fuel, watching it fail to turn over, and concluding the engine itself is useless, is close to the mistake a great deal of the biochar literature has made.
This argument must not be allowed to become impossible to disprove, and it is not. It does not claim every no-effect result is due to bad preparation; some soils are already fertile and buffered and have little to gain, which the reviews show plainly. The claim is narrower and testable: that charged, aged, seeded char will outperform fresh char, that the gap will widen over years rather than seasons, and that the comparison that would settle it, fresh versus charged versus charged-and-seeded, tracked over time against an explicit liming control, has rarely been run. That same comparison is what separates the biological claim from the chemistry, which is why the experiment in Section 10 carries the weight of the paper. The mixed record is not evidence against the idea. It is evidence that the decisive experiment has not yet been done.
9. Running lean: the thrift that falls out of the mechanism
If the earlier sections are right, the same arrangement that makes the soil last also makes it run lean, and it predicts that thrift as one thing falling out of the mechanism rather than a handful of separate savings bolted on. A community that has crossed into the self-feeding valley (Section 4) is, by its very nature, one that holds onto, recycles, and digs out its scarce resources instead of losing them, because that holding-on is exactly what keeps the valley deep. So efficiency is not an extra feature of the system; it is the system’s expected fingerprint. Each saving below is offered as a prediction whose size the experiment in Section 10 is built to measure, not as something already shown.
Water. Water thrift follows straight from the holding power of Section 2. The skeleton, and the living crumb structure it builds, soak up water and catch the flow that would otherwise drain past the roots, cutting evaporation and drainage and carrying the community through dry spells. There is a floor to this, set by the plant and not the soil, because a crop still has to run water through itself to pull carbon from the air; so the biggest gains show up in coarse, low-grip soils that had the least holding power to begin with.
Nutrients. Nutrient thrift follows from the same surface and the life it houses. Microbes lock nutrients into their own bodies and fungi ferry them back to the plant, recycling them inside the system, while the charged skeleton grips mineral nitrogen and other nutrients against leaching and against the losses that turn nitrogen back into gas. The prediction is a lower resupply rate, not a closed loop: what the harvest carries off still has to be put back, and the difference between a system that recycles tightly and a store that merely sits there is exactly the lasting-under-harvest tell of Section 8.
Nitrogen from the air, and a tuned engine. Two more savings are properly biological. The system can make some of its own nitrogen, through nitrogen-fixing microbes working alongside the plant-and-fungus network, an input limited not by how much nitrogen is in the air, which is most of it, but by getting that nitrogen to the fixers and shielding their machinery from oxygen, which a well-aired, well-aggregated topsoil is built to do. And the plant’s own metabolism runs closer to its ceiling once a complete set of minerals restores the trace-element parts its photosynthetic and respiratory machinery depend on, with the gain over worn-out ground large precisely because worn-out ground runs so far below that ceiling. None of these savings conjures resources from nothing; each converts and conserves what is supplied, so the system runs lean on its fuel, and the measure of how lean is the falling upkeep curve of the experiment in Section 10.
10. The one experiment that would settle it
The earlier sections show that every part the mechanism needs is independently established, and that five fields agree on its blueprint. What they do not show is the whole result: that a real soil can be driven across a threshold into a self-sustaining, terra-preta-like state. The synthesis is the groundwork; the demonstration would be the discovery. The design that follows is built to tell this paper's specific claim apart from the strongest rival, because it is not enough to show that a matured soil holds high function when you stop feeding it. A passive carbon-and-grip bank, the chemistry account granted in Section 8, predicts exactly that: a soil given durable carbon and base nutrients holds its gains for a long time simply because the carbon does not decay and the grip does not let go. The living-state claim makes a stronger, different prediction, and the experiment must isolate it.
The deciding variable is not stopping the inputs but continuing the harvest. A passive bank, however large, is drawn down by repeated cropping: each harvest removes nutrients, and with no resupply the store declines on a schedule set by its size. A soil whose fertility is kept up by a self-renewing community predicts the opposite, high function sustained across repeated harvests, because the community keeps weathering minerals, holding mobile nutrients, and rebuilding the pools faster than a passive store could. So the decisive contrast is performance under sustained cropping with outside inputs withheld, run long enough for a passive bank to draw down measurably. Lasting under harvest, not lasting at rest, is the signature of a living state.
The design is a single side-by-side field experiment on a degraded, low-carbon, low-grip soil of the kind where the mechanism matters most, with a real terra preta sample as a reference endpoint where available. Every plot gets the same total minerals and is brought to the same acidity and nutrient saturation, so that the carbon skeleton and the biology, not nutrients or acidity, are what is being tested. The plots are: (A) the full treatment, conditioned charcoal aged to high grip and mineral-loaded, plus a defined, locally boosted starter culture, at an over-the-threshold dose; (B) the same full package at a below-threshold dose; (C) a no-carbon plot getting the identical minerals and starter with no charcoal skeleton, optionally with an inert high-surface filler to separate physical structure from charcoal; (D) a correction control brought to the same acidity and nutrient status by liming and soluble fertilizer alone, without the skeleton and without the boosted starter, representing conventional practice; and (E) a bare-charcoal plot getting fresh, unconditioned char with no mineral loading or starter. An untreated degraded-soil baseline completes the set.
The thing measured is the whole terra preta package together, not yield alone, because the central claim is that the biological properties emerge as a set from one material rather than being installed one at a time. Across the cropping cycles each plot is scored for crop performance, microbial density and diversity and network complexity, disease suppression and signs of primed plant immunity, crumb stability and tilth, water-holding, and nutrient retention. The whole set emerging together in the full plot, appearing over time from material and starter without each function being engineered separately, is the positive signature of emergence; if the properties only appear when installed one by one, that part of the claim fails.
The contrasts decide the question. If the full plot (A) keeps high function under continued harvest while the below-threshold plot (B) slides back toward baseline, the result is a history-dependent alternative stable state with stickiness, two outcomes from the same endpoint conditions. If the full plot holds while the no-carbon plot (C) fails to establish or fails to keep the biological payoff, carbon is shown to be the keystone rather than one contributor among equals. If the full plot holds under harvest while the correction control (D) decays as its soluble store is cropped away, the result is shown to be more than a pH-and-nutrient fix, the very ground on which the chemistry account was granted its force. If the full plot beats the bare-charcoal plot (E), the mineral-loading and biological charging of the skeleton are shown to matter. The physical payoff, by contrast, is expected in every charcoal-bearing plot right away; it is not what the experiment is testing.
The cross-field picture also sharpens how to attempt the crossing, not just how to test it. Transferring suppressive soil shows the target community can be moved as a starter, and who-gets-there-first theory shows timing is decisive: the desired community should be established early enough, and densely enough, to claim the space before the degraded community reasserts itself. That the endpoint can be built by hand is not in doubt, since high-fertility dark earth has been made repeatedly by traditional practice in West Africa and the Amazon, a proof that the endpoint is reachable even if not that the controlled, instrumented, engineered route works. What has not been done is the demonstration of that route with the internal controls above.
Two supporting measurements turn specific claims into numbers within the same design. Tracking the forms phosphorus takes over time would show whether the system parks phosphorus in re-releasable forms or loses it to locked-away ones, and a drip-through column would test the anti-leaching claim directly. Varying the amount of weathered clay across otherwise identical full-treatment plots would locate the point where a clay surface flips from a re-chargeable phosphorus holder into a net trap that locks up both phosphorus and the microbial acids that would otherwise free it, the knife-edge that decides whether a phosphorus-grabbing material is an asset or a liability in a biologically intense soil.
The design is built to be disprovable in each of its claims, which is its purpose. A full plot that decays under continued harvest at the rate of the correction control would refute the living-state claim and vindicate the passive-bank account. A no-carbon plot that matches the full treatment would refute the keystone claim. A correction control that reproduces the whole package and holds it under cropping would show that liming and fertilizer are enough and that neither the skeleton nor the biology does the work the paper assigns them. The one thing the design does not allow is a free pass for a failed result: because the controls are built in and the plots differ one factor at a time, a failure of the full plot cannot be waved away as bad char or weak starter without that same excuse being tested against the other charcoal-bearing plots.
11. Conclusion
Terra preta is best understood not as a fertilized soil but as a soil whose microbial community has settled into a different equilibrium, and the proposed reason for that equilibrium is a durable, sponge-like charcoal skeleton acting as a shared, multi-purpose home. The skeleton supplies housing, nutrient grip, reservoirs, signal-holding, and shelter through different pore sizes and surface chemistries that ripen on different clocks; the resulting density and closeness switch on an efficiency economy run by crowd-sensing and a division of labor that pays only when partners are packed close, with gene-shedding deepening that economy over a longer evolutionary clock; the self-reinforcing loops of that economy are the formal requirement for an alternative stable state, a kind of state a growing body of evidence says soil communities can occupy; and the fungal network ties the system together, moves its minerals, trades back and forth with its hosts, and primes plant immunity, with all of these strengthening as the network densifies. The same organization also predicts that such a soil runs lean, holding water, recycling its nutrients, and renewing much of its own fertility under continued harvest, as one emergent consequence of the mechanism rather than a set of separate tricks. Five independent fields describe pieces of the same blueprint, and that agreement is the strongest support available now, though it motivates the idea rather than proving it.
The honest verdict is that the synthesis is strong, defensible, and testable, that its parts are individually well established, and that the whole result, a deliberate crossing into a self-sustaining high-fertility state with the tell-tale stickiness, is the open experiment that would tell a compelling idea apart from a proven discovery. The careful, calibrated version of the claim is also the more powerful one, because the corrections this synthesis builds in (a tipping point rather than a runaway, an efficient route rather than the only route, staggered ripening rather than instant function, measurable rather than conscious fungal signaling) are exactly what make the result survive scrutiny.
A short note on what is solid and what is proposed
In keeping with the technical paper: the individual parts of this mechanism are well established, the agreement of five fields on one blueprint is real, and the living dark earths of West Africa show the endpoint can be made by hand. What is proposed, not proven, is that these parts assemble into a single self-sustaining state in a soil that you can deliberately push a soil across that threshold. Two things this paper deliberately does not claim: that the fungal network is intelligent or communicates in the way popular accounts suggest, and that the centuries-long survival of dark earth is by itself proof of a living, self-renewing state (durable carbon alone would last that long). The one experiment that would settle it is the side-by-side test of Section 10, whose deciding measure is whether high fertility holds up under continued harvest.
This is the plain-language companion to the technical paper "Biochar: the Black 'Magic' of Terra Preta." For the scientific sources behind every claim above, and for the precise wording of the calibrated claims, see the technical paper.